Mikrotubule stanowią jeden z kluczowych elementów szkieletu komórkowego, bez którego życie komórkowe w znanej nam formie nie mogłoby istnieć. Ich obecność umożliwia zachowanie kształtu komórek, precyzyjne dzielenie materiału genetycznego, a także transport wewnątrzkomórkowy na znaczne – w skali komórki – odległości. Zrozumienie budowy i funkcji mikrotubul jest niezbędne nie tylko dla biologów komórki, lecz także dla medycyny, farmakologii i inżynierii biomedycznej, ponieważ liczne leki, choroby i procesy rozwojowe są z nimi ściśle powiązane.
Budowa mikrotubuli na poziomie molekularnym
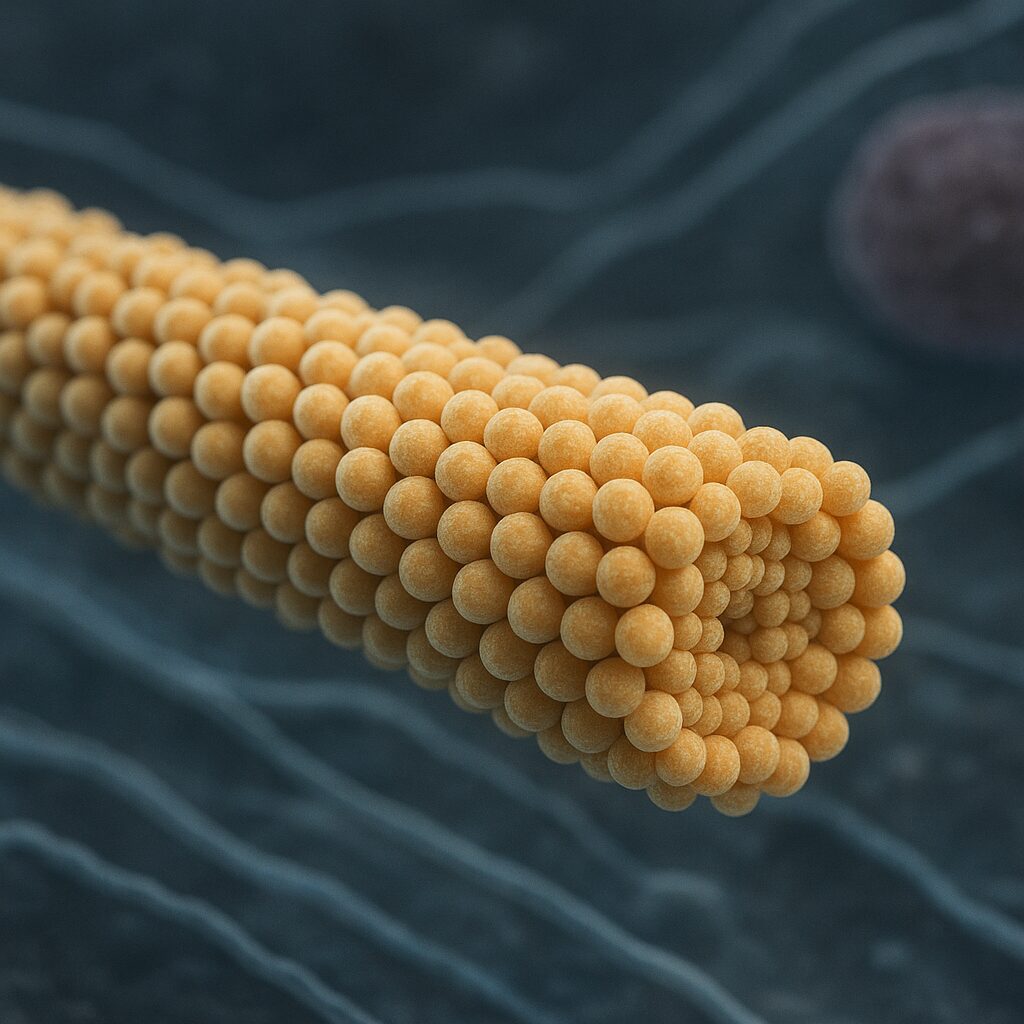
Mikrotubula jest długim, pustym w środku cylindrem zbudowanym z białek zwanych tubulinami. Jej ścianę tworzy zwykle 13 równolegle ułożonych protofilamentów, z których każdy jest liniowym łańcuchem dimerów tubuliny. Dimer ten składa się z dwóch ściśle ze sobą związanych podjednostek: alfa‑tubuliny i beta‑tubuliny. Obie należą do rodziny białek globularnych i wiążą nukleotyd GTP, jednak w odmienny sposób i z odmiennymi konsekwencjami dla dynamiki mikrotubuli.
Podjednostka alfa‑tubuliny wiąże cząsteczkę GTP, która pozostaje w niej trwale uwięziona i nie podlega wymianie w czasie funkcjonowania mikrotubuli. W przeciwieństwie do niej, beta‑tubulina wiąże GTP w sposób odwracalny – nukleotyd ten może być hydrolizowany do GDP, a następnie wymieniany na nową cząsteczkę GTP po odłączeniu dimeru od mikrotubuli. Właśnie ta różnica stanowi podstawę zjawiska zwanego dynamiką niestabilności, czyli naprzemiennych faz wzrostu i skracania mikrotubuli.
Mikrotubule są strukturami spolaryzowanymi. Oznacza to, że ich dwa końce nie są równoważne ani pod względem budowy, ani aktywności. Koniec określany jako koniec plus (lub dodatni) rośnie zwykle szybciej i jest bardziej dynamiczny, podczas gdy koniec minus (ujemny) jest względnie stabilny i zakotwiczony w określonych strukturach komórkowych. Ta polarność jest niezbędna do prawidłowego funkcjonowania białek motorycznych i do uporządkowania przestrzennego cytoszkieletu.
Średnica zewnętrzna mikrotubuli wynosi około 25 nm, a średnica wewnętrzna ok. 15 nm. Długość może sięgać od kilkuset nanometrów do wielu mikrometrów, a w komórkach nerwowych nawet dziesiątek mikrometrów. Pomimo wysokiej sztywności w porównaniu z innymi filamentami cytoszkieletu, mikrotubule są wciąż wystarczająco elastyczne, aby dopasowywać się do zmieniającego się środowiska komórkowego i uczestniczyć w dynamicznej reorganizacji wnętrza komórki.
Kluczową rolę w powstawaniu mikrotubul odgrywają kompleksy γ‑tubuliny, w tym tzw. γ‑TURC (gamma‑tubulin ring complex). Tworzą one pierścieniowe rusztowanie, które działa jak „matryca” inicjująca polimeryzację protofilamentów. W ten sposób komórka kontroluje liczbę, orientację i miejsce pojawiania się nowych mikrotubul, co ma znaczenie podczas podziałów komórkowych, migracji czy różnicowania.
Dynamika mikrotubuli i jej regulacja
Jedną z najbardziej fascynujących cech mikrotubul jest ich niezwykła dynamika. Polimeryzacja, czyli przyłączanie kolejnych dimerów tubuliny GTP do końca plus, odbywa się stosunkowo szybko, podobnie jak depolimeryzacja, polegająca na odłączaniu dimerów z beta‑tubuliną związaną z GDP. Zmiany te zachodzą spontanicznie, ale są precyzyjnie kontrolowane przez komórkę, co pozwala na błyskawiczne dostosowanie architektury cytoszkieletu.
Kluczowym zjawiskiem jest wspomniana niestabilność dynamiczna. Polega ona na naprzemiennych fazach wzrostu (rescue) i gwałtownego skracania (catastrophe) mikrotubuli. Gdy na końcu plus utrzymuje się tzw. czapeczka GTP – warstwa dimerów, w których beta‑tubulina nadal zawiera GTP – struktura jest stabilna i rośnie. Po utracie czapeczki, kiedy większość końcowych dimerów jest w formie GDP, mikrotubula staje się podatna na nagłe depolimeryzacje. To właśnie przejście od stanu wzrostu do stanu gwałtownego skracania określa się mianem katastrofy.
Komórka dysponuje rozbudowanym zestawem białek regulujących te procesy. Do najważniejszych należą białka MAP (microtubule‑associated proteins). Niektóre MAP, jak np. rodziny MAP2 czy Tau, stabilizują mikrotubule, wiążąc się wzdłuż ich powierzchni i ograniczając częstotliwość katastrof. Inne, takie jak katanina, mogą rozcinać mikrotubule, co prowadzi do szybkiej reorganizacji sieci cytoszkieletu. Istnieją również białka, które specyficznie rozpoznają rosnące końce plus – tzw. +TIPs – i kierują je w odpowiednie regiony komórki.
Dynamika mikrotubul jest ściśle powiązana z cyklem komórkowym. W fazie interfazowej sieć mikrotubul organizuje cytoplazmę, pozycjonuje organella i wspiera transport. Jednak w momencie wejścia w mitozę większość mikrotubul ulega depolimeryzacji, a następnie powstaje nowa, silnie zorganizowana struktura – wrzeciono podziałowe, odpowiedzialne za rozdział chromosomów. Tak daleko posunięta przebudowa cytoszkieletu w tak krótkim czasie byłaby niemożliwa bez zjawiska niestabilności dynamicznej.
Na dynamikę mikrotubul wpływają także warunki fizykochemiczne, takie jak stężenie tubuliny, obecność jonów magnezu, temperatura oraz czynniki chemiczne, w tym liczne związki o działaniu farmakologicznym lub toksycznym. Niektóre z nich, jak taksany, stabilizują mikrotubule, uniemożliwiając ich depolimeryzację, inne, jak kolchicyna czy alkaloidy z zimowitu, hamują polimeryzację. W obu przypadkach dochodzi do zaburzeń funkcji komórki, co może zostać wykorzystane terapeutycznie, zwłaszcza w onkologii.
Rola mikrotubuli w komórce: rusztowanie, transport i podział
Jedną z najbardziej podstawowych funkcji mikrotubuli jest utrzymywanie kształtu komórki oraz zapewnienie jej mechanicznej odporności. W komórkach zwierzęcych sieć mikrotubul promieniście rozchodzi się od centrosomu w kierunku obwodu, pełniąc rolę wewnętrznego „rusztowania”. Umożliwia to komórce wytrzymywanie sił ściskających i rozciągających, a także zachowanie spójności strukturalnej podczas ruchu czy zmian objętości.
Niemniej istotna jest rola mikrotubuli w organizacji rozmieszczenia organelli. Aparat Golgiego, lizosomy, endosomy czy mitochondria lokalizują się w określonych regionach komórki właśnie dzięki interakcji z siecią mikrotubul. Zmiana organizacji mikrotubuli może prowadzić do przemieszczenia organelli, co obserwuje się m.in. w czasie różnicowania komórek nabłonkowych, powstawania neuronów czy w procesach sekrecyjnych.
Jednak prawdopodobnie najbardziej spektakularną funkcją mikrotubuli jest udział w transporcie wewnątrzkomórkowym. Po powierzchni mikrotubul poruszają się białka motoryczne, takie jak kinezyny i dyneiny. Wykorzystując energię z hydrolizy ATP, przemieszczają one pęcherzyki, kompleksy białkowe, mRNA, a nawet całe organella wzdłuż wyznaczonych tras. Kinezyny transportują zazwyczaj ładunek w kierunku końca plus, natomiast dyneiny – w stronę końca minus. Ta dwukierunkowość ruchu jest fundamentalna dla prawidłowego funkcjonowania złożonych komórek, zwłaszcza neuronów.
W komórkach nerwowych mikrotubule układają się równolegle w aksonach i dendrytach, tworząc swoiste „autostrady” dla pęcherzyków synaptycznych, receptorów i elementów cytoplazmy. Zaburzenia w ich organizacji lub w funkcjonowaniu białek motorycznych mogą prowadzić do poważnych deficytów transportu aksonalnego, co wiąże się z neurodegeneracją, osłabieniem przewodzenia impulsów oraz stopniową utratą funkcji neuronalnych.
Nie można pominąć kluczowego znaczenia mikrotubuli w podziale komórkowym. Podczas mitozy i mejozy tworzą one wrzeciono podziałowe, którego zadaniem jest równomierny rozdział chromosomów do komórek potomnych. Mikrotubule kinetochorowe przyczepiają się do specjalnych struktur na chromosomach – kinetochorów – i za pomocą skoordynowanych procesów depolimeryzacji oraz pracy białek motorycznych przeciągają chromatydy siostrzane do przeciwległych biegunów komórki. Równolegle mikrotubule biegunowe oraz astralne stabilizują wrzeciono i pozycjonują je względem błony komórkowej.
Zaburzenia w funkcjonowaniu mikrotubuli podczas podziału mogą prowadzić do nieprawidłowej segregacji chromosomów, aneuploidii i w konsekwencji nowotworzenia. Dlatego tak ważna jest precyzja mechanizmów regulujących długość, stabilność i interakcje mikrotubuli w trakcie mitozy. Komórka wykorzystuje w tym celu liczne białka checkpointowe, które monitorują przyłączenie mikrotubuli do kinetochorów oraz napięcie generowane między chromosomami.
Mikrotubule w rzęskach i wici: aparat ruchowy komórki
Specjalną kategorię struktur opartych na mikrotubulach stanowią rzęski i wici, określane wspólnie jako wyspecjalizowane wypustki komórkowe. Ich wewnętrzny szkielet – aksonema – ma charakterystyczny układ 9+2: dziewięć par mikrotubul obwodowych otacza centralną parę. W niektórych typach rzęsek, np. w pierwotnych rzęskach czuciowych, brakuje centralnej pary, a organizacja przypomina układ 9+0.
Ruch rzęsek i wici jest wynikiem skoordynowanej pracy dynein aksonemalnych, które ślizgają się po sąsiednich mikrotubulach, generując siłę zginającą całą strukturę. Dzięki temu rzęski nabłonka oddechowego mogą przemieszczać warstwę śluzu, usuwając zanieczyszczenia i mikroorganizmy z dróg oddechowych, a wici plemników zapewniają ich zdolność do poruszania się w kierunku komórki jajowej.
Nie wszystkie rzęski pełnią jednak funkcję ruchową. Tzw. rzęski pierwotne są najczęściej nieruchome i pełnią rolę anten czuciowych, zbierając informacje o otoczeniu chemicznym i mechanicznym. Uczestniczą w kluczowych ścieżkach sygnałowych, takich jak hedgehog, Wnt czy PDGF. Zaburzenia w budowie lub funkcjonowaniu rzęsek, określane jako ciliopatie, prowadzą do szerokiego spektrum chorób, obejmujących wady rozwojowe, schorzenia nerek, wątroby, siatkówki czy defekty rozrodu.
U organizmów jednokomórkowych, takich jak pierwotniaki, rzęski i wici są często podstawowym narzędziem lokomocji i zdobywania pokarmu. Pozwalają na aktywne przemieszczanie się w środowisku wodnym, kierunkowe reagowanie na bodźce świetlne, chemiczne czy grawitacyjne, a także na tworzenie prądów wodnych ułatwiających pobieranie cząstek pokarmowych. Ewolucyjna konserwacja budowy aksonemy wskazuje, że mikrotubule stanowiły bardzo wcześnie fundament aparatów ruchu komórkowego.
Centrosom, ciałka podstawne i organizacja mikrotubuli
Mikrotubule nie powstają chaotycznie w dowolnych miejscach cytoplazmy. Ich organizację w komórkach zwierzęcych koordynują wyspecjalizowane struktury nazywane centrami organizacji mikrotubuli (MTOC). Najbardziej znanym MTOC jest centrosom, zlokalizowany zwykle w pobliżu jądra komórkowego. Składa się on z dwóch centrioli ułożonych pod kątem prostym oraz otaczającej je pericentriolowej macierzy białkowej.
Centriola ma postać cylindrycznej struktury zbudowanej z dziewięciu tripletów mikrotubul, ułożonych w charakterystyczny pierścień. O ile mikrotubule tworzące trzon komórki są dynamiczne, o tyle tripletowa struktura centrioli jest stabilna i pełni głównie funkcję organizacyjną. Z pericentriolowej macierzy wyrastają liczne mikrotubule cytoplazmatyczne, których końce minus są tam zakotwiczone, podczas gdy końce plus rosną w kierunku obwodu komórki. W czasie podziału komórkowego centrosom duplikuje się, tworząc dwa bieguny wrzeciona mitotycznego.
Blisko spokrewnioną strukturą z centriolą jest ciałko podstawne (bazalne), które stanowi punkt wyjścia dla rzęski lub wici. Pod względem ultrastruktury ciałko podstawne przypomina centriolę, ale jego głównym zadaniem jest zakotwiczenie i organizacja mikrotubul aksonemy. W wielu komórkach ta sama centriola, która pełniła funkcję elementu centrosomu, może przekształcić się w ciałko podstawne rzęski, co podkreśla płynność relacji między tymi organellami.
W komórkach roślinnych brak typowego centrosomu z centriolami, a mimo to mikrotubule są doskonale zorganizowane. Funkcję MTOC pełnią wówczas rejony związane z jądrem komórkowym, błoną plazmatyczną lub aparatem Golgiego. Rośliny wytworzyły więc alternatywne mechanizmy organizacji mikrotubuli, co dowodzi, że ewolucja cytoszkieletu może przebiegać różnymi drogami, przy zachowaniu zasadniczych właściwości polimerów tubulinowych.
Znaczenie kliniczne mikrotubuli i leki mikrotubuloaktywne
Rola mikrotubuli wykracza daleko poza czysto teoretyczne aspekty biologii komórki. Stanowią one istotny cel wielu leków stosowanych w terapii chorób nowotworowych, pasożytniczych, a także w badaniach nad neurodegeneracją. Leki te, nazywane mikrotubuloaktywnymi, modyfikują dynamikę polimeryzacji tubuliny, co prowadzi do zahamowania podziałów komórkowych lub innych kluczowych funkcji.
Do najważniejszych substancji stabilizujących mikrotubule należy paklitaksel i inne taksany. Wiążą się one z beta‑tubuliną wewnątrz mikrotubuli, wzmacniając jej strukturę i ograniczając depolimeryzację. Efektem jest „zamrożenie” wrzeciona mitotycznego w nieprawidłowej konfiguracji, co uniemożliwia prawidłowy rozdział chromosomów i prowadzi do śmierci szybko dzielących się komórek nowotworowych. Z kolei alkaloidy barwinka, takie jak winkrystyna czy winblastyna, hamują polimeryzację i promują depolimeryzację, również destabilizując wrzeciono.
W terapii przeciwpasożytniczej wykorzystuje się związki z grupy benzimidazoli, które selektywnie zakłócają funkcjonowanie mikrotubuli pasożytów, w mniejszym stopniu wpływając na komórki gospodarza. Upośledzają one procesy wchłaniania składników odżywczych i podział komórek pasożyta, co prowadzi do jego obumarcia. Przykładem jest albendazol czy mebendazol, szeroko stosowane w leczeniu zakażeń nicieniami.
Mikrotubule odgrywają również dużą rolę w patogenezie chorób neurodegeneracyjnych. Białko Tau, należące do klasy białek MAP, stabilizuje mikrotubule w aksonach neuronów. W chorobie Alzheimera i pokrewnych tauopatiach ulega ono nieprawidłowym modyfikacjom potranslacyjnym, szczególnie hiperfosforylacji, co obniża jego powinowactwo do mikrotubuli. Skutkiem jest destabilizacja sieci mikrotubul, zaburzenie transportu aksonalnego i gromadzenie wewnątrzkomórkowych agregatów Tau – splątków neurofibrylarnych. Proces ten przyczynia się do stopniowego obumierania neuronów.
Innym przykładem są neuropatie obwodowe wywołane toksynami środowiskowymi lub działaniem niektórych leków przeciwnowotworowych. Nadmierne stabilizowanie lub destabilizowanie mikrotubuli w neuronach może upośledzać transport aksonalny, prowadząc do drętwienia, bólu, osłabienia mięśni czy zaburzeń czucia. Zrozumienie mechanizmów leżących u podstaw tych powikłań jest kluczowe dla opracowywania terapii o większej selektywności i mniejszej toksyczności neurologicznej.
Ewolucja, różnorodność i przyszłe kierunki badań nad mikrotubulami
Mikrotubule są niezwykle konserwatywnymi ewolucyjnie strukturami. Białka tubulinowe wykazują wysoki stopień podobieństwa od jednokomórkowych eukariontów po organizmy wielokomórkowe, co sugeruje, że pojawiły się bardzo wcześnie w historii życia. Jednocześnie istnieje bogactwo izotypów tubuliny, różniących się nieznacznie sekwencją aminokwasową i ulegających specyficznym modyfikacjom potranslacyjnym, takim jak acylacja, tyrozynacja, detyrozynacja czy glutamylacja.
Zróżnicowanie to określa się jako „kod tubulinowy”, analogicznie do „kodu histonowego” w chromatynie. Uważa się, że kombinacje izotypów i modyfikacji tubulin determinują właściwości poszczególnych mikrotubul, ich podatność na działanie białek motorycznych i MAP, a także ich długość życia i funkcję w komórce. Odkrycie tego kodu i jego roli w organizmach żywych jest jednym z fascynujących wyzwań współczesnej biologii komórki.
Nowoczesne techniki mikroskopii superrozdzielczej, kriomikroskopii elektronowej i obrazowania in vivo pozwalają śledzić zachowanie pojedynczych mikrotubul w żywych komórkach z niespotykaną dotąd dokładnością. Badania te ujawniają subtelne zróżnicowanie dynamiki w różnych regionach komórki, interakcje z błonami, kompleksami białkowymi oraz wpływ lokalnych sygnałów molekularnych. Dzięki nim powstają coraz bardziej realistyczne modele komputerowe zachowania całej sieci cytoszkieletu.
Istotnym kierunkiem badań jest również wykorzystanie mikrotubul i białek motorycznych w inżynierii biomolekularnej. Tworzy się układy hybrydowe, w których mikrotubule stanowią tory dla sztucznie zaprojektowanych kinezyn przenoszących nanocząstki, ładunki leków czy elementy elektroniczne. Projektuje się też mikroskopijne „silniki” oparte na motoryce dynein i kinezyn, które w przyszłości mogą znaleźć zastosowanie w mikromaszynach medycznych.
Wreszcie, coraz większe znaczenie ma badanie mikrotubul w kontekście mechanobiologii. Komórki odczuwają i przetwarzają bodźce mechaniczne, takie jak rozciąganie, ucisk czy sztywność podłoża. Mikrotubule, wraz z innymi komponentami cytoszkieletu, uczestniczą w transdukcji tych bodźców do jąder komórkowych i wpływają na ekspresję genów. Zrozumienie tych zależności pomoże wyjaśnić procesy różnicowania komórek macierzystych, powstawania nowotworów czy przebudowy tkanek pod wpływem treningu fizycznego.
Najczęściej zadawane pytania (FAQ)
Co to jest mikrotubula i z czego się składa?
Mikrotubula to cylindryczny element cytoszkieletu o średnicy około 25 nm, zbudowany z powtarzających się dimerów białkowych alfa‑ i beta‑tubuliny. Dimery te tworzą równoległe protofilamenty, których zwykle jest trzynaście, ułożonych w pustą w środku rurkę. Struktura ta jest spolaryzowana – posiada koniec plus i koniec minus – co umożliwia kierunkowy transport wewnątrzkomórkowy oraz dynamiczne wydłużanie i skracanie w odpowiedzi na potrzeby komórki.
Jaką rolę mikrotubule pełnią w podziale komórki?
Podczas podziału komórkowego mikrotubule tworzą wrzeciono podziałowe, które odpowiada za równomierne rozdzielenie chromosomów do komórek potomnych. Mikrotubule kinetochorowe przyłączają się do specjalnych struktur na chromosomach i, skracając się oraz współpracując z białkami motorycznymi, „przeciągają” chromatydy w stronę biegunów komórki. Mikrotubule biegunowe i astralne stabilizują wrzeciono i pozycjonują je tak, by płaszczyzna podziału przebiegała we właściwym miejscu.
W jaki sposób mikrotubule biorą udział w transporcie wewnątrzkomórkowym?
Mikrotubule pełnią rolę szlaków, po których poruszają się białka motoryczne – głównie kinezyny i dyneiny. Kinezyny zwykle transportują ładunek w stronę końca plus, czyli ku obwodowi komórki, natomiast dyneiny kierują się do końca minus, zlokalizowanego zwykle w pobliżu centrosomu. Do tych białek przyłączone są pęcherzyki, organella czy kompleksy białkowe, które dzięki energii z hydrolizy ATP mogą być szybko i precyzyjnie przemieszczane na duże, jak na skalę komórki, odległości.
Dlaczego mikrotubule są celem działania wielu leków przeciwnowotworowych?
Komórki nowotworowe dzielą się bardzo intensywnie, dlatego są szczególnie wrażliwe na zaburzenia powstawania wrzeciona mitotycznego. Leki takie jak taksany czy alkaloidy barwinka modyfikują dynamikę mikrotubuli: jedne nadmiernie je stabilizują, inne hamują polimeryzację. W obu przypadkach dochodzi do nieprawidłowego rozdziału chromosomów i aktywacji mechanizmów prowadzących do śmierci komórki. Dzięki temu można selektywniej niszczyć populacje szybko proliferujących komórek guza.
Jakie choroby mogą wynikać z nieprawidłowości w budowie lub funkcji mikrotubuli?
Defekty mikrotubuli i białek z nimi związanych prowadzą do szerokiego spektrum schorzeń. Należą do nich m.in. tauopatie, w tym choroba Alzheimera, w których zaburzona jest stabilizacja mikrotubuli w neuronach, co upośledza transport aksonalny. Ciliopatie, wynikające z nieprawidłowej budowy rzęsek, powodują wady rozwojowe, choroby nerek, zaburzenia płodności i schorzenia siatkówki. Mikrotubule biorą też udział w powstawaniu nowotworów poprzez wpływ na prawidłową segregację chromosomów podczas podziałów komórkowych.

