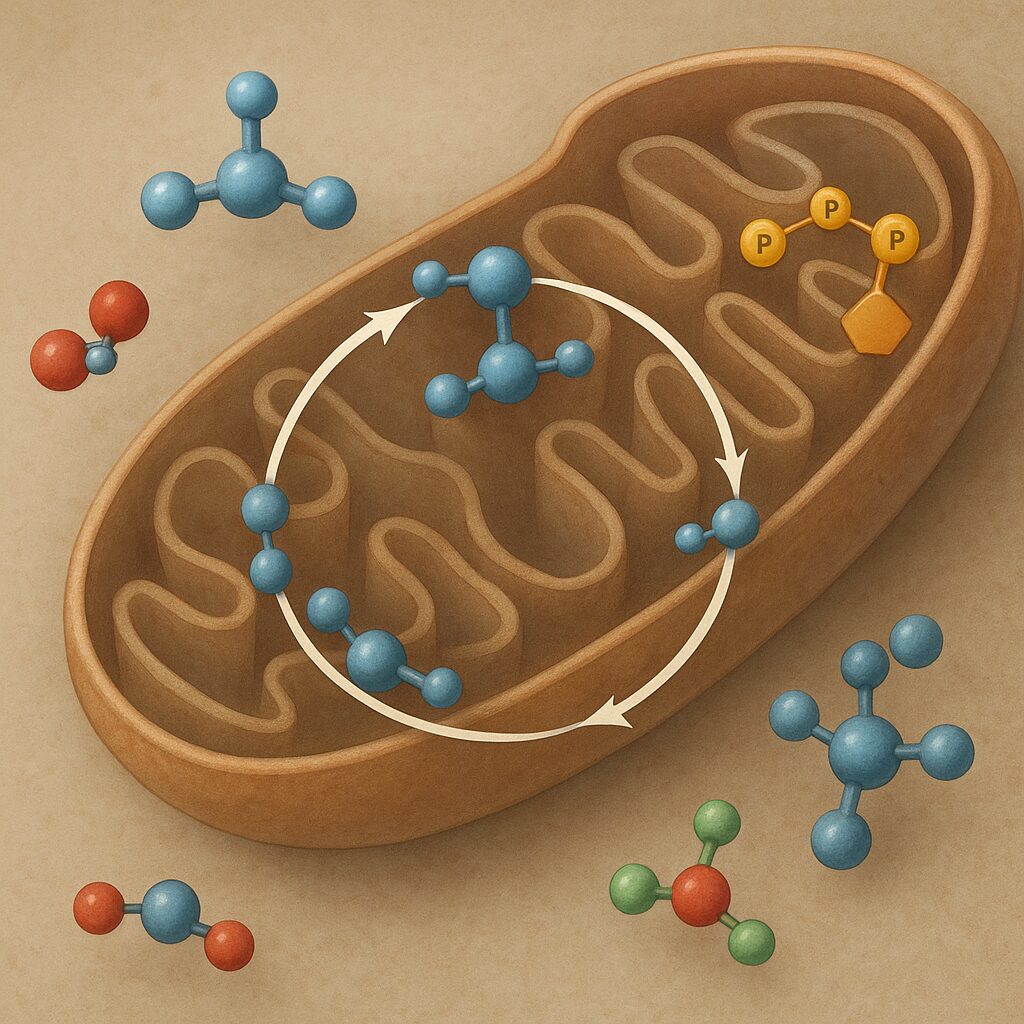
Cykl Krebsa, nazywany także cyklem kwasu cytrynowego lub cyklem kwasów trójkarboksylowych, jest jednym z najważniejszych szlaków metabolicznych w komórkach organizmów tlenowych. Odgrywa kluczową rolę w pozyskiwaniu energii z pokarmu, łącząc procesy rozkładu węglowodanów, tłuszczów i białek w jeden spójny mechanizm biochemiczny. Zrozumienie jego przebiegu pomaga wyjaśnić, jak komórki przekształcają energię chemiczną zawartą w związkach organicznych w uniwersalną walutę energetyczną organizmu – cząsteczki ATP.
Historia odkrycia i znaczenie cyklu Krebsa
Badania nad przemianami metabolicznymi w komórkach rozpoczęły się intensywnie na przełomie XIX i XX wieku, kiedy rozwijała się biochemia. Jednak dopiero prace Hansa Adolfa Krebsa, niemieckiego biochemika pracującego w Wielkiej Brytanii, doprowadziły do zidentyfikowania pełnego cyklu reakcji zachodzących w mitochondriach. W 1937 roku Krebs opisał sekwencję przemian, w której związki dwuwęglowe są stopniowo utleniane z wytworzeniem dwutlenku węgla, zredukowanych koenzymów i energii. Za to odkrycie otrzymał Nagrodę Nobla w 1953 roku.
Odkrycie cyklu Krebsa było przełomem, ponieważ po raz pierwszy połączono wiele pojedynczych reakcji w jeden spójny schemat, wyjaśniający, w jaki sposób komórki pozyskują energię z różnych źródeł. Co istotne, cykl ten nie jest drogą jednokierunkową służącą wyłącznie do rozkładu cząsteczek. Spełnia także funkcje biosyntetyczne, dostarczając prekursorów do syntezy aminokwasów, porfiryn, kwasów tłuszczowych czy glukozy. Stanowi zatem centralny węzeł metaboliczny, do którego dochodzą różne szlaki i z którego odgałęzia się wiele innych procesów.
Znaczenie cyklu Krebsa wykracza poza samą biochemię komórki. Jego prawidłowe działanie warunkuje sprawne funkcjonowanie całego organizmu. Zaburzenia w enzymach cyklu prowadzą do poważnych chorób metabolicznych, problemów z układem nerwowym, niewydolności mięśni czy zaburzeń rozwojowych u dzieci. Dlatego cykl kwasu cytrynowego jest intensywnie badany zarówno w kontekście podstawowych procesów życiowych, jak i w medycynie, farmakologii czy naukach o żywieniu.
Budowa i lokalizacja cyklu Krebsa w komórce
Cykl Krebsa zachodzi w strukturach komórkowych zwanych mitochondria. Są one często określane mianem „elektrowni komórkowych”, ponieważ tu zachodzi większość procesów związanych z wytwarzaniem ATP. U organizmów eukariotycznych mitochondria posiadają podwójną błonę – zewnętrzną, stosunkowo gładką i przepuszczalną dla wielu małych cząsteczek, oraz wewnętrzną, silnie pofałdowaną, tworzącą grzebienie mitochondrialne. Właśnie w macierzy mitochondrialnej zlokalizowane są enzymy cyklu kwasu cytrynowego.
Substratem wyjściowym cyklu jest acetylo-CoA – związek powstający głównie z glukoza w procesie glikolizy i dalszego przekształcenia pirogronianu, ale również z kwasów tłuszczowych w wyniku β-oksydacji oraz z niektórych aminokwasów w czasie ich katabolizmu. Acetylo-CoA wprowadza do cyklu dwuwęglową jednostkę, która następnie ulega stopniowemu utlenieniu do CO₂. Podczas tych przemian powstają zredukowane formy koenzymów NADH i FADH₂, a także jedna cząsteczka wysokoenergetycznego nukleotydu GTP (lub ATP, w zależności od typu komórki).
Przebieg cyklu Krebsa można podzielić na kilka kluczowych etapów:
- kondensacja acetylo-CoA ze szczawiooctanem i powstanie cytrynianu,
- izomeryzacja cytrynianu do izocytrynianu,
- kolejne etapy dekarboksylacji oksydacyjnej, generujące NADH i CO₂,
- utlenianie bursztynianu do fumaranu z udziałem FAD,
- uwodnienie fumaranu do jabłczanu,
- utlenianie jabłczanu do szczawiooctanu, który zamyka cykl.
Istotne jest to, że sam cykl nie jest bezpośrednio głównym źródłem ATP. Powstaje w nim stosunkowo niewielka ilość GTP/ATP, natomiast zasadniczy zysk energetyczny pochodzi z późniejszego utleniania NADH i FADH₂ w łańcuchu oddechowym, zlokalizowanym w wewnętrznej błonie mitochondrialnej. To tam energia uwolniona z przenoszenia elektronów na tlen jest wykorzystywana do syntezy dużych ilości ATP w procesie fosforylacji oksydacyjnej.
Etapy reakcji w cyklu kwasu cytrynowego
Choć szczegółowy opis wszystkich reakcji cyklu Krebsa może wydawać się skomplikowany, zrozumienie ich kolejności i funkcji pozwala dostrzec elegancję tego szlaku. Każda reakcja jest katalizowana przez wyspecjalizowany enzym, a całość przebiega w sposób cykliczny, co oznacza, że związek wejściowy – szczawiooctan – jest w końcowym etapie odtwarzany.
Powstanie cytrynianu i przejście do izocytrynianu
Pierwszą reakcją cyklu jest kondensacja acetylo-CoA (dwu atomy węgla) ze szczawiooctanem (cztery atomy węgla), katalizowana przez syntazę cytrynianową. Powstaje sześcioatomowy cytrynian, a cząsteczka CoA zostaje uwolniona. Jest to etap silnie egzoergiczny, co sprawia, że praktycznie nieodwracalny w warunkach fizjologicznych i pełni funkcję jednej z głównych reakcji regulacyjnych cyklu.
Następnie cytrynian jest przekształcany w izocytrynian przy udziale akonitazy. Reakcja ta zachodzi dwustopniowo – najpierw powstaje nietrwały cis-akonitan, a następnie następuje jego uwodnienie do izocytrynianu. Zmiana konfiguracji przestrzennej przygotowuje cząsteczkę do kolejnych etapów utleniania i dekarboksylacji.
Dekarboksylacje oksydacyjne i powstawanie NADH
Kolejnym ważnym etapem jest utlenianie izocytrynianu do alfa-ketoglutaranu. Reakcja ta, katalizowana przez dehydrogenazę izocytrynianową, obejmuje jednoczesne usunięcie grupy karboksylowej w postaci CO₂ oraz redukcję NAD⁺ do NADH. Alfa-ketoglutaran jest pięciowęglowym związkiem pośrednim, kluczowym również dla syntezy niektórych aminokwasów, np. glutaminianu.
Następnie alfa-ketoglutaran ulega kolejnej dekarboksylacji oksydacyjnej, prowadzącej do powstania bursztynylo-CoA. Za tę reakcję odpowiada kompleks dehydrogenazy alfa-ketoglutaranowej, działający mechanistycznie podobnie do kompleksu dehydrogenazy pirogronianowej. W jej wyniku powstaje druga cząsteczka CO₂ i kolejna cząsteczka NADH. Bursztynylo-CoA, zawierający wysokoenergetyczne wiązanie tioestrowe, stanowi bezpośrednie źródło energii do syntezy GTP lub ATP.
Powstawanie GTP/ATP i redukcja FAD
Przekształcenie bursztynylo-CoA w bursztynian jest katalizowane przez syntetazę bursztynylo-CoA. Energia zgromadzona w wiązaniu tioestrowym zostaje wykorzystana do fosforylacji GDP do GTP (lub ADP do ATP, zależnie od typu komórki i izoenzymu). To jedyny etap cyklu Krebsa, w którym bezpośrednio powstaje wysokoenergetyczny nukleotyd trifosforanowy. Bursztynian jest czterowęglowym kwasem dikarboksylowym, który podlega dalszemu utlenianiu.
Dalsza reakcja polega na utlenieniu bursztynianu do fumaranu przez dehydrogenazę bursztynianową. W przeciwieństwie do dehydrogenaz wykorzystujących NAD⁺, tutaj koenzymem jest FAD, redukowany do FADH₂. Enzym ten jest częścią kompleksu II łańcucha oddechowego, co oznacza, że elektrony z bursztynianu są bezpośrednio wprowadzane do systemu transportu elektronów, omijając pierwszy kompleks, który przyjmuje elektrony z NADH.
Odtworzenie szczawiooctanu i zamknięcie cyklu
Fumaran ulega uwodnieniu do jabłczanu w reakcji katalizowanej przez fumarazę. To stosunkowo prosta przemiana, polegająca na dodaniu cząsteczki wody do wiązania podwójnego. Następnie jabłczan jest utleniany do szczawiooctanu przez dehydrogenazę jabłczanową, czemu towarzyszy redukcja NAD⁺ do NADH. Powstały szczawiooctan może ponownie reagować z acetylo-CoA, co rozpoczyna kolejny obrót cyklu. W ten sposób cykl Krebsa jest układem samonapędzającym się, pod warunkiem, że dostępna jest odpowiednia ilość substratu wejściowego i tlenu jako końcowego akceptora elektronów w łańcuchu oddechowym.
Znaczenie cyklu Krebsa dla metabolizmu komórki
Cykl kwasu cytrynowego jest często porównywany do centralnego węzła, od którego odchodzą liczne drogi metaboliczne. Kluczowym aspektem jest jego rola w pozyskiwaniu energii. W trakcie jednego obrotu cyklu, z jednej cząsteczki acetylo-CoA powstają trzy cząsteczki NADH, jedna cząsteczka FADH₂ oraz jedna cząsteczka GTP/ATP. Po uwzględnieniu udziału tych zredukowanych koenzymów w łańcuchu oddechowym otrzymujemy znaczny zysk energetyczny, który jest niezbędny do podtrzymania procesów życiowych komórki.
Rola cyklu Krebsa nie ogranicza się jednak wyłącznie do katabolizmu. Wiele jego pośrednich produktów pełni funkcje biosyntetyczne. Alfa-ketoglutaran stanowi prekursor glutaminianu i innych aminokwasów, bursztynylo-CoA uczestniczy w syntezie porfiryn, w tym hemu w hemoglobinie i cytochromach, a szczawiooctan jest punktem wyjścia do glukoneogenezy, czyli syntezy glukozy z niecukrowych substratów. Dzięki temu cykl kwasu cytrynowego jest jednocześnie szlakiem katabolicznym i anabolicznym – jest szlakiem amfiboliczny.
Ważnym zjawiskiem są także reakcje anaplerotyczne, czyli procesy uzupełniające puli związków pośrednich cyklu, gdy są one zużywane do biosyntezy. Przykładem jest karboksylacja pirogronianu do szczawiooctanu, katalizowana przez karboksylazę pirogronianową. Bez takich reakcji cykl uległby „rozładowaniu”, ponieważ kolejne cząsteczki pośrednie byłyby wyciągane do innych szlaków bez odpowiedniego uzupełniania. Reakcje anaplerotyczne zapewniają zatem stabilność działania cyklu.
Cykl Krebsa ściśle współpracuje z innymi elementami metabolizmu komórki, takimi jak łańcuch oddechowy, glikoliza, β-oksydacja kwasów tłuszczowych czy szlaki biosyntezy aminokwasów. Ta wzajemna integracja umożliwia elastyczne dostosowanie przemian metabolicznych do aktualnych potrzeb energetycznych i biosyntetycznych komórki. W warunkach wysokiego zapotrzebowania na ATP przyspiesza tempo przepływu przez cykl, natomiast w sytuacjach, gdy potrzebne są głównie prekursory biosyntetyczne, część związków pośrednich jest kierowana w stronę szlaków anabolicznych.
Regulacja cyklu Krebsa i czynniki wpływające na jego aktywność
Dla komórki kluczowe jest precyzyjne regulowanie tempa, z jakim zachodzi cykl kwasu cytrynowego. Zbyt niska aktywność prowadziłaby do niedoboru ATP i zredukowanych koenzymów, natomiast nadmierne tempo mogłoby powodować niepotrzebne zużycie substratów i zaburzenia równowagi metabolicznej. Regulacja odbywa się na kilku poziomach, obejmując dostępność substratów, stężenie produktów, oddziaływania allosteryczne oraz modyfikacje enzymów.
Najważniejszym czynnikiem sterującym jest stan energetyczny komórki. Wysoki poziom ATP hamuje kluczowe enzymy cyklu, takie jak dehydrogenaza izocytrynianowa czy kompleks dehydrogenazy alfa-ketoglutaranowej. Z kolei wzrost stężenia ADP i AMP, będących wskaźnikami niskiej energii, stymuluje ich aktywność. Podobnie NADH, będący produktem reakcji utleniania, hamuje enzymy dehydrogenazowe, co zapobiega nadmiernemu gromadzeniu zredukowanych koenzymów, gdy łańcuch oddechowy pracuje zbyt wolno.
Dodatkowo na przebieg cyklu wpływa dostępność tlenu, który jest ostatecznym akceptorem elektronów w łańcuchu oddechowym. W warunkach niedotlenienia transport elektronów zostaje spowolniony, co prowadzi do nagromadzenia NADH i FADH₂. To z kolei hamuje enzymy cyklu zależne od NAD⁺ i FAD, zmniejszając tempo jego działania. Komórka przechodzi wtedy na mniej efektywne, ale niezależne od tlenu szlaki, takie jak fermentacja mlekowa w mięśniach.
Regulację cyklu Krebsa wzmacniają także mechanizmy hormonalne, szczególnie w komórkach tkanek zwierzęcych. Hormony regulujące gospodarkę energetyczną, takie jak adrenalina, glukagon czy insulina, wpływają pośrednio na dostępność substratów, tempo glikolizy, β-oksydacji i glukoneogenezy, a tym samym na ilość acetylo-CoA wprowadzanej do cyklu. W tkankach o wysokim zapotrzebowaniu energetycznym, np. w mięśniach szkieletowych czy sercu, istnieją dodatkowe mechanizmy dostosowujące aktywność enzymów do intensywności pracy.
Cykl Krebsa w zdrowiu i chorobie
Ze względu na centralną rolę w metabolizmie, cykl kwasu cytrynowego jest wrażliwy na różnego rodzaju zaburzenia genetyczne, toksyny, niedobory witamin czy stres oksydacyjny. Wiele enzymów cyklu wymaga obecności kofaktorów pochodzących z witamin, takich jak tiamina (witamina B₁), ryboflawina (B₂), niacyna (B₃) czy kwas pantotenowy (B₅). Ich brak może zaburzać działanie poszczególnych reakcji, prowadząc do kumulacji pośrednich metabolitów i obniżenia produkcji ATP.
Wrodzone defekty enzymów cyklu Krebsa, choć rzadkie, są zazwyczaj bardzo poważne. Mogą prowadzić do encefalopatii, opóźnienia rozwoju psychoruchowego, kwasicy metabolicznej, problemów z funkcjonowaniem mięśni czy serca. Przykładem jest niedobór dehydrogenazy alfa-ketoglutaranowej, który powoduje zaburzenia w przemianie aminokwasów i poważne deficyty energetyczne. Leczenie takich schorzeń bywa trudne i często koncentruje się na łagodzeniu objawów oraz unikania czynników pogarszających metabolizm.
Cykl Krebsa odgrywa też istotną rolę w patogenezie chorób nowotworowych. Komórki nowotworowe często przeprogramowują metabolizm, korzystając w dużym stopniu z glikolizy beztlenowej, nawet przy obecności tlenu (efekt Warburga). Nie oznacza to jednak całkowitego wyłączenia cyklu Krebsa, lecz raczej jego przebudowę pod kątem dostarczania prekursorów biosyntetycznych niezbędnych do szybkich podziałów. Zrozumienie tych zmian jest podstawą badań nad nowymi terapiami onkologicznymi, które celują w specyficzne różnice metaboliczne między komórkami zdrowymi a nowotworowymi.
Innym obszarem, w którym cykl kwasu cytrynowego ma duże znaczenie, są choroby neurodegeneracyjne i proces starzenia. Mitochondria neuronów są szczególnie wrażliwe na uszkodzenia oksydacyjne i zaburzenia w transporcie elektronów. Spadek sprawności cyklu Krebsa oraz łańcucha oddechowego może prowadzić do niedoboru ATP, zwiększonej produkcji reaktywnych form tlenu i ostatecznie do śmierci komórek nerwowych. Badania nad modulowaniem aktywności mitochondrialnej oraz ochroną enzymów cyklu są ważnym kierunkiem w poszukiwaniu terapii choroby Alzheimera, Parkinsona czy stwardnienia zanikowego bocznego.
Cykl Krebsa w kontekście ewolucji i różnorodności organizmów
Choć cykl kwasu cytrynowego jest najlepiej poznany u organizmów tlenowych, jego elementy i warianty występują również wśród bakterii i archeonów, które żyją w bardzo zróżnicowanych warunkach środowiskowych. Niektóre mikroorganizmy korzystają z odwróconej wersji cyklu, tzw. cyklu rewersyjnego, w którym dwutlenek węgla jest wiązany i przekształcany w związki organiczne. Stanowi to alternatywną ścieżkę autotroficznej asymilacji węgla, niezależną od cyklu Calvina.
Różnice w budowie i funkcjonowaniu cyklu Krebsa między organizmami odzwierciedlają ich adaptacje do określonych nisz ekologicznych i dostępnych źródeł energii. U niektórych bakterii brak jest pełnego zestawu enzymów, co prowadzi do niekompletnych cykli, które jednak wciąż pełnią ważne funkcje biosyntetyczne. Z kolei organizmy roślinne posiadają mitochondria podobne do zwierzęcych, ale ich cykl Krebsa współdziała z procesami fotosyntezy zachodzącymi w chloroplastach, tworząc skomplikowaną sieć powiązań metabolicznych.
Z perspektywy ewolucyjnej obecność bardzo podobnego cyklu u tak wielu różnych organizmów sugeruje, że powstał on na wczesnym etapie rozwoju życia na Ziemi i był na tyle efektywny, że został zachowany w toku dalszej ewolucji. Modyfikacje, które zaszły w poszczególnych liniach rodowych, pokazują, jak szlaki metaboliczne mogą być dostosowywane do zmieniających się warunków, zachowując jednocześnie swoją podstawową funkcję – zapewnienie efektywnego obiegu węgla i energii w komórce.
Praktyczne zastosowania wiedzy o cyklu Krebsa
Znajomość cyklu kwasu cytrynowego ma znaczenie nie tylko w badaniach podstawowych, ale również w wielu dziedzinach praktycznych. W medycynie zrozumienie przebiegu cyklu i jego regulacji pozwala lepiej interpretować wyniki badań laboratoryjnych, zwłaszcza dotyczących metabolizmu wątroby, mięśni czy serca. Nieprawidłowe stężenia niektórych metabolitów mogą wskazywać na zaburzenia w konkretnych etapach cyklu lub w powiązanych szlakach, co pomaga w diagnostyce chorób metabolicznych.
W farmakologii enzymy cyklu Krebsa stanowią potencjalne cele dla leków przeciwnowotworowych, przeciwbakteryjnych czy przeciwpasożytniczych. Ponieważ komórki patogenów często mają zmienione lub specyficzne warianty enzymów, możliwe jest opracowywanie związków hamujących ich aktywność bez silnego oddziaływania na enzymy gospodarza. Dotyczy to zwłaszcza mikroorganizmów chorobotwórczych, które wykorzystują cykl Krebsa w warunkach niszowych, np. w środowisku o ograniczonej dostępności tlenu.
W dietetyce i naukach o żywieniu wiedza o cyklu kwasu cytrynowego pozwala lepiej zrozumieć, jak organizm wykorzystuje różne makroskładniki – węglowodany, tłuszcze i białka – jako źródła energii. W zależności od rodzaju diety i poziomu aktywności fizycznej zmienia się stosunek udziału poszczególnych substratów w tworzeniu acetylo-CoA. U sportowców wytrzymałościowych adaptacje mitochondrialne prowadzą do zwiększenia aktywności enzymów cyklu oraz łańcucha oddechowego, co przekłada się na większą wydolność i lepsze wykorzystanie kwasów tłuszczowych jako paliwa.
W biotechnologii i inżynierii metabolicznej modyfikacja cyklu Krebsa i jego połączeń z innymi szlakami umożliwia projektowanie mikroorganizmów o zwiększonej wydajności produkcji określonych metabolitów, np. kwasów organicznych, aminokwasów czy związków użytecznych w przemyśle chemicznym. Manipulowanie przepływem węgla przez cykl, poprzez nadekspresję lub wyciszanie konkretnych enzymów, jest jednym z kluczowych narzędzi w projektowaniu komórek fabryk.
Najważniejsze pojęcia związane z cyklem Krebsa
Aby uporządkować wiedzę o cyklu kwasu cytrynowego, warto wyróżnić kilka kluczowych pojęć, które pomagają w dalszym zgłębianiu tematu:
- mitochondria – organelle komórkowe będące głównym miejscem wytwarzania energii w postaci ATP,
- acetylo-CoA – wspólny produkt rozkładu węglowodanów, tłuszczów i części białek, wprowadzany do cyklu,
- ATP – podstawowa cząsteczka przenosząca energię w komórce,
- NADH i FADH₂ – zredukowane koenzymy przenoszące elektrony do łańcucha oddechowego,
- łańcuch oddechowy – zespół kompleksów białkowych w błonie mitochondrialnej, odpowiedzialny za fosforylację oksydacyjną,
- reakcje anaplerotyczne – procesy uzupełniające pulę pośrednich związków cyklu,
- glukoza – podstawowy cukier, główne źródło energii w wielu komórkach,
- amfiboliczny – określenie szlaku pełniącego zarówno funkcje kataboliczne, jak i anaboliczne,
- β-oksydacja – proces rozkładu kwasów tłuszczowych prowadzący do powstawania acetylo-CoA,
- glukoneogeneza – szlak syntezy glukozy z niecukrowych prekursorów, powiązany z cyklem Krebsa.
Zrozumienie tych pojęć i relacji między nimi jest podstawą do dalszego zgłębiania bardziej zaawansowanych zagadnień biochemicznych. Cykl kwasu cytrynowego pozostaje jednym z najlepiej zbadanych, a jednocześnie wciąż fascynujących szlaków metabolicznych, ilustrujących złożoność i harmonijność przemian zachodzących w komórkach wszystkich organizmów tlenowych.
FAQ – najczęściej zadawane pytania o cykl Krebsa
Dlaczego cykl Krebsa jest tak ważny dla organizmu?
Cykl Krebsa to centralny szlak metaboliczny, w którym ostatecznie spotykają się losy większości węglowodanów, tłuszczów i części białek. W jego trakcie powstają zredukowane koenzymy NADH i FADH₂, przekazujące elektrony do łańcucha oddechowego, gdzie syntetyzowane są duże ilości ATP. Jednocześnie pośrednie produkty cyklu służą jako prekursory do biosyntezy aminokwasów, hemu czy glukozy, dlatego szlak ten łączy funkcje energetyczne i budulcowe.
Gdzie w komórce zachodzi cykl Krebsa i jakie struktury biorą w nim udział?
Cykl kwasu cytrynowego zachodzi w macierzy mitochondrialnej, czyli wewnętrznej przestrzeni mitochondriów otoczonej błoną wewnętrzną. Tam zlokalizowane są enzymy katalizujące kolejne etapy cyklu. Zredukowane koenzymy NADH i FADH₂ powstałe w cyklu oddają elektrony do łańcucha oddechowego w błonie wewnętrznej mitochondrium. Współdziałanie macierzy i błony wewnętrznej umożliwia sprzężenie utleniania substratów z produkcją ATP.
Jak cykl Krebsa łączy się z innymi szlakami metabolicznymi?
Cykl Krebsa jest połączony z glikolizą, β-oksydacją i katabolizmem aminokwasów poprzez acetylo-CoA, który staje się substratem wejściowym cyklu. Z kolei jego pośrednie produkty, takie jak alfa-ketoglutaran, bursztynylo-CoA czy szczawiooctan, są wykorzystywane w syntezie aminokwasów, hemu oraz w glukoneogenezie. Dzięki temu cykl działa jak centrum przesiadkowe metabolizmu, umożliwiając płynne przekierowywanie węgla między rozkładem a biosyntezą.
Czy cykl Krebsa może działać bez obecności tlenu?
Same reakcje cyklu Krebsa nie wymagają bezpośrednio tlenu, ale jego ciągła praca zależy od sprawnego działania łańcucha oddechowego, w którym tlen jest końcowym akceptorem elektronów. Jeśli brakuje tlenu, gromadzi się NADH i FADH₂, a ich utlenianie zostaje zahamowane. Wysokie stężenie tych zredukowanych koenzymów hamuje dehydrogenazy cyklu, co spowalnia lub zatrzymuje jego działanie. Dlatego w warunkach beztlenowych komórki przechodzą na mniej wydajne szlaki, np. fermentację.
Jak dieta i aktywność fizyczna wpływają na cykl Krebsa?
Dieta dostarcza substratów, które po przekształceniu do acetylo-CoA wchodzą do cyklu Krebsa. Wysokie spożycie węglowodanów zwiększa udział glukozy, natomiast dieta bogata w tłuszcze nasila β-oksydację i wykorzystanie kwasów tłuszczowych. Regularna aktywność fizyczna zwiększa liczbę mitochondriów w mięśniach oraz aktywność enzymów cyklu i łańcucha oddechowego. Dzięki temu wzrasta zdolność do produkcji ATP i wykorzystania kwasów tłuszczowych jako źródła energii podczas wysiłku.

