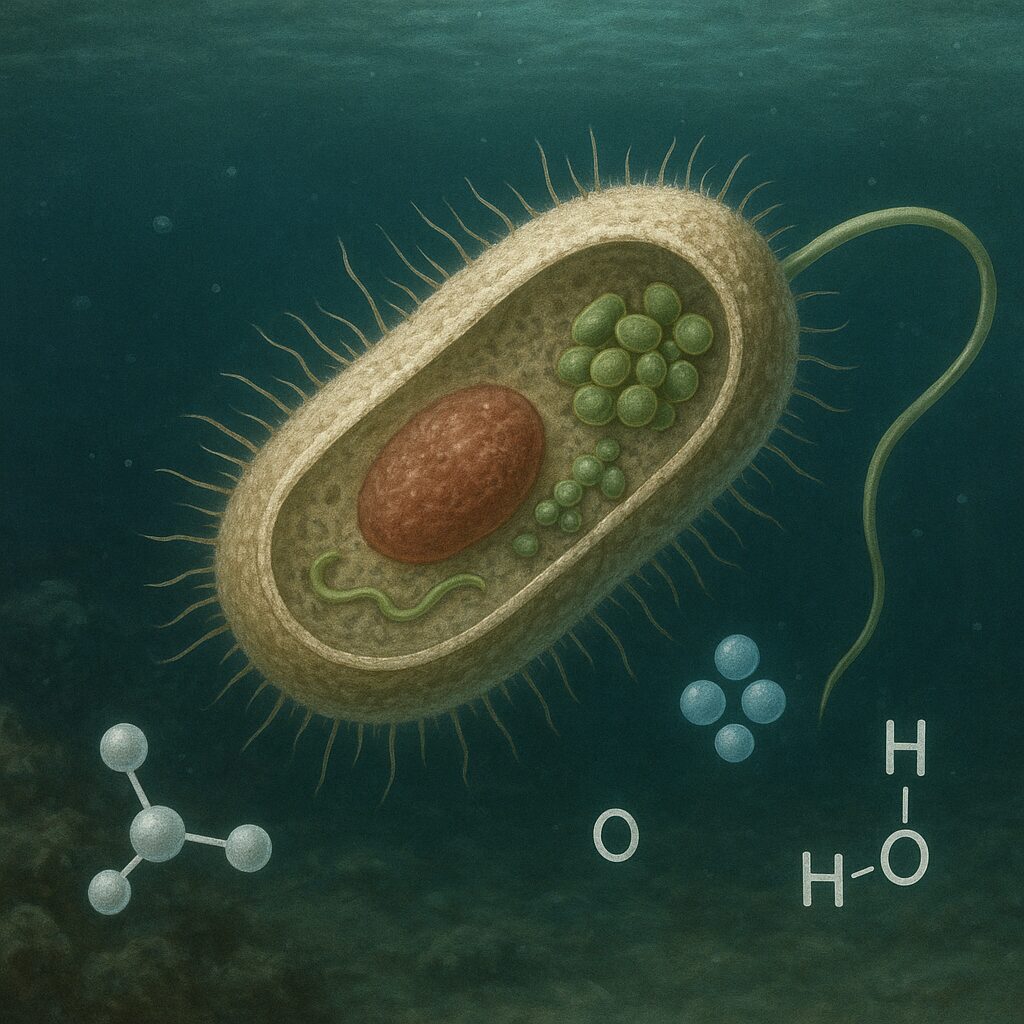
Chemoautotrofy stanowią jedną z najbardziej fascynujących i niedocenianych grup organizmów na Ziemi. W przeciwieństwie do znanych wszystkim roślin zielonych nie potrzebują światła słonecznego, by wytwarzać własną materię organiczną. Wykorzystują energię z reakcji chemicznych, dzięki czemu mogą zasiedlać środowiska całkowicie pozbawione światła, takie jak głębiny oceaniczne czy wnętrza skał. Zrozumienie ich funkcjonowania pozwala lepiej poznać nie tylko biosferę naszej planety, ale także potencjalne warunki życia poza Ziemią.
Podstawowa definicja i miejsce chemoautotrofów w świecie organizmów
Termin chemoautotrof składa się z trzech członów: „chemo–” oznacza, że energia pochodzi z reakcji chemicznych, „auto–” wskazuje na samodzielność w wytwarzaniu związków organicznych, a „–trof” odnosi się do sposobu odżywiania. Chemoautotrofy to zatem organizmy, które jako źródło energii wykorzystują reakcje utleniania nieorganicznych związków chemicznych, a jako źródło węgla używają dwutlenku węgla (CO₂), przekształcając go w związki organiczne w procesie autotrofii.
W klasycznej biologii wyróżnia się kilka głównych typów strategii odżywiania:
- fotoautotrofy – czerpią energię ze światła (rośliny, sinice, część protistów);
- chemoautotrofy – energię pozyskują z reakcji chemicznych utleniania związków nieorganicznych;
- heterotrofy – pobierają gotową materię organiczną (zwierzęta, grzyby, większość bakterii);
- miksyotrofy – łączą różne strategie (np. niektóre protisty).
Chemoautotrofy zalicza się głównie do domen Bacteria i Archaea. To przede wszystkim bakterie siarkowe, żelazowe, nitryfikacyjne, a także liczne archeony żyjące w skrajnych warunkach. Ich wspólną cechą jest zdolność do chemolitoautotrofii, czyli wykorzystywania nieorganicznych donorów elektronów (np. jonów amonowych, siarkowodoru, żelaza(II), wodoru) do syntezy ATP, a następnie do asymilacji CO₂.
W ujęciu ekologicznym chemoautotrofy są producentami pierwotnymi w środowiskach, gdzie światło w ogóle nie dociera. Tworzą podstawę łańcuchów pokarmowych w takich miejscach jak kominy hydrotermalne na dnie oceanów czy głębokie warstwy skorupy ziemskiej. Stanowią tym samym dowód, że życie nie jest nierozerwalnie związane z energią słoneczną.
Mechanizmy pozyskiwania energii i asymilacji dwutlenku węgla
Funkcjonowanie chemoautotrofów opiera się na dwóch zasadniczych etapach: pozyskiwaniu energii z nieorganicznych reakcji redoks oraz wykorzystaniu tej energii do wiązania węgla w postaci CO₂ w cząsteczki organiczne. Oba procesy są ze sobą ściśle skorelowane i wymagają wyspecjalizowanych układów enzymatycznych.
Reakcje chemiczne jako źródło energii
Chemoautotrofy wykorzystują energię z reakcji utleniania związków nieorganicznych. Przykładowe pary donor–akceptor elektronów to:
- jon amonowy (NH₄⁺) → azotyn (NO₂⁻) lub azotan (NO₃⁻);
- siarkowodór (H₂S) → siarka elementarna (S) lub siarczany (SO₄²⁻);
- żelazo(II) Fe²⁺ → żelazo(III) Fe³⁺;
- wodór cząsteczkowy (H₂) → protony (H⁺);
- tlenek węgla (CO) → dwutlenek węgla (CO₂).
W tych reakcjach dochodzi do przepływu elektronów przez łańcuch transportu elektronów w błonie komórkowej, co prowadzi do wytworzenia gradientu protonowego. Następnie syntaza ATP wykorzystuje ten gradient do produkcji cząsteczek ATP. Energia zawarta w ATP, a także siła redukcyjna w postaci NADH czy NADPH, służy później do redukcji CO₂.
Istotne jest, że proces ten nie wymaga fotonów. Można go porównać do oddychania komórkowego, z tą różnicą, że donorami elektronów są proste związki nieorganiczne, a organizm nie musi zużywać związków organicznych. W wielu środowiskach to chemiczna energia Ziemi, nie promieniowanie słoneczne, napędza całą lokalną biosferę.
Szlaki asymilacji dwutlenku węgla
Podobnie jak rośliny, liczne chemoautotrofy wykorzystują cykl Calvina-Bensona do włączania CO₂ w związki organiczne. Enzymem kluczowym jest RuBisCO, który katalizuje karboksylację rybulozo-1,5-bisfosforanu. Produktami pośrednimi cyklu są m.in. triozy, z których mogą powstawać cukry, aminokwasy i inne związki niezbędne do budowy komórki.
Jednak wiele archeonów i niektóre bakterie wykorzystują alternatywne szlaki, dostosowane do warunków środowiskowych i dostępnych donorów energii. Przykłady to:
- cykl odwrotnej redukcji kwasu trikarboksylowego (odwrotny cykl Krebsa);
- cykl hydroksypropionianowy i hydroksybutyrylowy;
- szlak Wooda–Ljungdahla (szlak acetylokoenzymu A), typowy dla części archeonów i bakterii acetogennych.
Te alternatywne mechanizmy często są bardziej energooszczędne niż klasyczny cykl Calvina, co ma kluczowe znaczenie w środowiskach o skrajnie niskiej dostępności energii. Dzięki temu chemoautotrofy potrafią przetrwać i rozwijać się w miejscach, gdzie zasoby energetyczne są minimalne lub niestabilne.
Bilans energetyczny i wydajność chemoautotrofii
Energia uzyskiwana z utleniania nieorganicznych związków jest zwykle mniejsza niż energia pozyskiwana przez rośliny z fotosyntezy, licząc na jednostkę zredukowanego węgla. Z tego powodu tempo wzrostu chemoautotrofów jest często niższe, a ich biomasa rzadziej osiąga tak duże wartości jak biomasa roślin lądowych czy fitoplanktonu.
Mimo to efektywność ekologiczna chemoautotrofów jest ogromna. Tam, gdzie występują stabilne źródła związków chemicznych (np. emisje siarkowodoru, amoniaku, wodoru), mogą one wytwarzać znaczące ilości biomasy, która staje się pokarmem dla organizmów heterotroficznych. Szczególnie wyraźnie widać to w ekosystemach związanych z aktywnością wulkaniczną i hydrotermalną.
Różnorodność taksonomiczna i przykłady chemoautotrofów
Chemoautotrofy nie stanowią jednej zwartej grupy systematycznej. Są rozproszone wśród wielu linii ewolucyjnych bakterii i archeonów, co sugeruje, że zdolność do chemoautotrofii pojawiała się wielokrotnie w dziejach życia lub była dziedziczona z bardzo dawnych, wspólnych przodków. Ich różnorodność przejawia się zarówno w typie wykorzystywanych związków chemicznych, jak i w preferowanych siedliskach.
Bakterie nitryfikacyjne
Bakterie nitryfikacyjne pełnią kluczową rolę w globalnym obiegu azotu. Zalicza się do nich dwie główne grupy funkcjonalne:
- bakterie utleniające amon (AOB – ammonia-oxidizing bacteria), np. z rodzaju Nitrosomonas;
- bakterie utleniające azotyny (NOB – nitrite-oxidizing bacteria), np. z rodzaju Nitrobacter.
Organizmy te utleniają jon amonowy do azotynów, a następnie azotyny do azotanów, czerpiąc z tych reakcji energię. W środowiskach glebowych, słodkowodnych i morskich odpowiadają za przekształcanie związków azotowych, co ma ogromne znaczenie dla dostępności azotu dla roślin i dla zapobiegania toksycznym stężeniom amoniaku.
Bakterie nitryfikacyjne są klasycznym przykładem chemoautotrofów: wykorzystują energię z utleniania związków azotu, a CO₂ służy im jako źródło węgla do budowy komórki. Proces nitryfikacji zachodzi najintensywniej w środowiskach dobrze natlenionych, takich jak gleby uprawne, strefa przypowierzchniowa osadów dennych czy biologiczne oczyszczalnie ścieków.
Bakterie siarkowe
Bakterie siarkowe wykorzystują zredukowane formy siarki, takie jak siarkowodór, tiosiarczany czy siarka elementarna, jako donory elektronów. Wśród nich wyróżnia się organizmy żyjące zarówno w warunkach tlenowych, jak i beztlenowych. Przykładem są rodzaje Thiobacillus czy Beggiatoa.
W środowiskach, gdzie zachodzi rozkład materii organicznej w warunkach beztlenowych (np. w osadach dennych, namułach, strefach beztlenowych jezior), powstaje siarkowodór. Gdy staje się on dostępny w strefach kontaktu z tlenem, bakterie siarkowe mogą go utleniać, pozyskując energię do asymilacji CO₂. Powstające siarczany mogą być następnie redukowane przez inne mikroorganizmy, co tworzy pełny cykl siarkowy.
Bakterie te przyczyniają się do neutralizowania toksycznego H₂S w ekosystemach, a także uczestniczą w procesach geochemicznych prowadzących do powstawania skał siarczanowych. Ich obecność ma również znaczenie technologiczne, np. w procesach bioutleniania rud metali.
Bakterie żelazowe i manganowe
Niektóre bakterie zdolne są do utleniania jonów żelaza(II) i manganu(II), co prowadzi do powstawania osadów tlenków tych metali. Organizmy takie jak Gallionella czy Leptothrix zasiedlają bogate w żelazo wody podziemne, źródła i strefy przejściowe między środowiskiem beztlenowym i tlenowym.
W tych mikrosiedliskach żelazo(II), rozpuszczalne w wodzie, zostaje utlenione do żelaza(III), które wytrąca się w postaci tlenków. Bakterie żelazowe wykorzystują tę reakcję jako źródło energii, a powstające osady mogą kształtować strukturę podłoża, blokować filtry studzienne czy wpływać na skład chemiczny wód. Podobne znaczenie mają mikroorganizmy utleniające mangan.
Archeony metanogenne i metanotroficzne
W domenie Archaea szczególnie interesujące są organizmy związane z przemianami metanu. Archeony metanogenne, choć głównie chemoautotroficzne lub chemomiksotroficzne, wytwarzają metan w procesach beztlenowych z prostych substratów, takich jak wodór i CO₂ czy octany. Z kolei część archeonów i bakterii metanotroficznych może wykorzystywać metan lub tlenek węgla jako źródło energii, jednocześnie asymilując CO₂ lub węgiel z utlenianego związku.
W niektórych głębinowych środowiskach, takich jak zimne wycieki metanowe, powstają całe zespoły organizmów opierających swój metabolizm na utlenianiu metanu lub siarkowodoru. Wiele z tych mikroorganizmów żyje w symbiozie z bezkręgowcami, przekazując im część wytworzonej biomasy i stanowiąc podstawę egzystencji całej społeczności.
Chemoautotrofy w ekstremalnych środowiskach
Chemoautotrofy odgrywają szczególnie ważną rolę w ekosystemach ekstremalnych. Należą do nich:
- kominy hydrotermalne na dnie oceanów, gdzie gorące, bogate w siarkę i metale roztwory wydobywają się z wnętrza Ziemi;
- zimne wycieki metanowe w głębokich basenach morskich;
- gorące źródła i gejzery bogate w siarkę;
- głębokie warstwy skorupy ziemskiej, w tym wody podziemne i pory skalne;
- kwaśne wody kopalniane zawierające wysokie stężenia metali i siarczanów.
W takich siedliskach światło słoneczne nie dociera lub warunki fizykochemiczne (temperatura, pH, zasolenie) są skrajne. Mimo to chemoautotrofy potrafią tam nie tylko przetrwać, ale też tworzyć gęste populacje. Wysoka różnorodność metaboliczna i odporność na stres środowiskowy czynią je jednymi z najbardziej przystosowanych form życia.
Znaczenie chemoautotrofów w ekosystemach i cyklach biogeochemicznych
Rola chemoautotrofów wykracza daleko poza ich niszowe siedliska. Biorą udział w obiegach pierwiastków, stabilizują chemizm środowiska i podtrzymują życie w miejscach pozbawionych światła. Ich aktywność wpływa na bilans gazów cieplarnianych, produktywność ekosystemów i procesy geologiczne, kształtując oblicze planety na skalach czasowych liczących miliony lat.
Udział w globalnych cyklach pierwiastków
Chemoautotrofy są kluczowymi uczestnikami obiegu takich pierwiastków jak azot, siarka, żelazo, mangan, węgiel i wodór. Przykładowo:
- w cyklu azotu przeprowadzają nitryfikację, przekształcając amon w azotany dostępne dla roślin;
- w cyklu siarki utleniają siarkowodór i inne formy zredukowane do siarczanów;
- w cyklu żelaza utleniają jony Fe²⁺, wpływając na osady żelaziste i jakość wód;
- w cyklu węgla mogą wykorzystywać CO₂ jako jedyne źródło węgla i wpływać na jego magazynowanie w biomase czy osadach.
Wiele reakcji biogeochemicznych, które dawniej przypisywano wyłącznie procesom abiotycznym, okazało się ściśle związanych z aktywnością mikroorganizmów chemoautotroficznych. Na przykład powstawanie masywnych złóż rudy żelaza czy siarczanów wiąże się z ich wielowiekowym metabolizmem.
Podstawy łańcuchów pokarmowych niezależnych od światła
Jednym z najbardziej spektakularnych przejawów znaczenia chemoautotrofów są ekosystemy kominów hydrotermalnych. Na głębokości kilku kilometrów pod powierzchnią oceanu, w całkowitej ciemności, wokół szczelin emitujących gorące płyny bogate w siarkę i metale, rozwijają się bogate w życie społeczności.
Podstawą tych ekosystemów są bakterie i archeony chemoautotroficzne, które utleniają siarkowodór lub metan, wytwarzając materię organiczną. Z chemoautotrofami wchodzą w symbiozę bezkręgowce, takie jak ruryczniki, małże czy ślimaki. Mikroorganizmy mieszkają w ich tkankach i dostarczają im substancji odżywczych, podczas gdy gospodarze zapewniają im dostęp do odpowiednich płynów i tlenu.
Takie ekosystemy dowodzą, że złożone społeczności biologiczne mogą istnieć bez udziału fotosyntezy, bazując wyłącznie na energii geochemicznej. Podobne układy stwierdzono również w głębiach lądowych, np. w kopalniach złota czy głębokich odwiertach, gdzie woda podziemna stale dostarcza substraty chemiczne.
Wpływ na klimat i geochemię oceanów
Aktywność chemoautotrofów może wpływać na bilans gazów cieplarnianych, zwłaszcza metanu i CO₂. Mikroorganizmy utleniające metan (metanotrofy) ograniczają jego emisję z osadów dennych do atmosfery, co ma znaczenie dla regulacji efektu cieplarnianego. Z kolei procesy związane z utlenianiem siarkowodoru w strefach beztlenowych oceanów mogą wpływać na rozkład tlenu i skład chemiczny wód.
W głębinach morskich chemoautotrofy przyczyniają się do wiązania niektórych pierwiastków śladowych w osadach, tworząc trwałe minerały. W ten sposób pośrednio regulują dostępność metali dla organizmów planktonowych i innych komponentów biosfery oceanicznej. Długoterminowe konsekwencje tych procesów odzwierciedlają się w zapisie geologicznym, np. w formie charakterystycznych warstw osadowych.
Funkcja w glebie i wodach słodkich
W ekosystemach lądowych chemoautotrofy obecne w glebie wpływają na żyzność i zdrowie roślin. Bakterie nitryfikacyjne przekładają amon, powstający m.in. z rozkładu materii organicznej i nawozów, na azotany. Z jednej strony zwiększa to dostępność azotu dla roślin, z drugiej może prowadzić do wypłukiwania azotanów do wód i do zjawiska eutrofizacji.
W wodach słodkich chemoautotrofy uczestniczą w samooczyszczaniu się zbiorników. Utlenianie zredukowanych form azotu i siarki ogranicza toksyczne działanie amoniaku i siarkowodoru na organizmy wodne. W osadach dennych utrzymują równowagę między strefami tlenowymi i beztlenowymi, wpływając na stabilność całego ekosystemu zbiornika.
Chemoautotrofia a początki życia i poszukiwanie życia pozaziemskiego
Pojęcie chemoautotrofii ma kluczowe znaczenie dla teorii dotyczących początku życia na Ziemi oraz dla astrobiologii. Wiele hipotez zakłada, że pierwsze organizmy żywe były organizmami chemolitoautotroficznymi, zależnymi od energii z reakcji zachodzących na powierzchni minerałów i w pobliżu kominów hydrotermalnych.
Hipotezy o hydrotermalnym pochodzeniu życia
Jedna z głównych hipotez głosi, że życie mogło powstać w mikrokieszeniach porowatych skał w pobliżu kominów hydrotermalnych. Tam gradienty temperatury i pH, a także obecność zredukowanych związków nieorganicznych, sprzyjały powstawaniu prostych układów metabolicznych opartych na reakcjach redoks. Pierwotne systemy mogły przypominać współczesne chemoautotrofy, wykorzystujące wodór, siarczki i dwutlenek węgla.
W takich warunkach minerały żelazowo-siarczkowe mogły pełnić rolę katalizatorów, ułatwiając powstawanie pierwszych związków organicznych. Z czasem mogły się rozwinąć bardziej złożone sieci reakcji, w tym prymitywne szlaki asymilacji CO₂. Chemoautotroficzny tryb życia mógł zatem poprzedzać pojawienie się fotosyntezy i tlenowego oddychania.
Znaczenie dla astrobiologii
Obserwacja, że życie może funkcjonować w całkowitej ciemności, bazując na chemoautotrofii, ma bezpośrednie implikacje dla poszukiwań życia pozaziemskiego. Ciała niebieskie takie jak Europa (księżyc Jowisza) czy Enceladus (księżyc Saturna) posiadają podlodowe oceany, prawdopodobnie w kontakcie z aktywną geologicznie skorupą skalną.
Jeżeli w takich oceanach zachodzą reakcje między wodą a skałami (np. serpentynizacja), mogą powstawać wodór, metan oraz inne zredukowane związki chemiczne. Stanowiłyby one potencjalne źródło energii dla hipotetycznych chemoautotrofów. Brak światła słonecznego nie musiałby więc oznaczać braku życia, o ile istnieją odpowiednie gradienty chemiczne.
Badania misji kosmicznych poszukują śladów aktywności geochemicznej oraz obecności związków takich jak wodór, siarkowodór czy metan w atmosferach i pióropuszach tych ciał. Wykrycie stabilnych gradientów chemicznych byłoby pośrednim dowodem na możliwość istnienia ekosystemów chemoautotroficznych w odległych zakątkach Układu Słonecznego.
Zastosowania praktyczne i znaczenie dla człowieka
Choć chemoautotrofy działają często poza zasięgiem ludzkiej percepcji, ich właściwości mogą być wykorzystywane w technologii, ochronie środowiska i rolnictwie. Zrozumienie ich metabolizmu jest istotne zarówno dla ograniczania negatywnych skutków działalności człowieka, jak i dla tworzenia nowatorskich bioprocesów.
Biotechnologia i oczyszczanie środowiska
Chemoautotrofy od dawna stosowane są w inżynierii środowiska. W oczyszczalniach ścieków bakterie nitryfikacyjne i siarkowe uczestniczą w usuwaniu związków azotu i siarki, zapobiegając eutrofizacji oraz emisji odorów. Kontrolowane warunki pozwalają na optymalizację ich aktywności i stabilne działanie całych instalacji.
Innym zastosowaniem jest bioutlenianie rud, w którym bakterie utleniają siarczki metali, ułatwiając odzysk cennych pierwiastków, takich jak miedź czy złoto. Ten proces może być mniej inwazyjny niż klasyczne metody metalurgiczne, choć wymaga kontroli, by ograniczać zakwaszanie środowiska i powstawanie kwaśnych wód kopalnianych.
Rolnictwo i zarządzanie glebą
W rolnictwie aktywność chemoautotrofów, zwłaszcza bakterii nitryfikacyjnych, jest czynnikiem warunkującym dostępność azotu. Świadome zarządzanie nawożeniem i warunkami glebowymi może wpływać na tempo nitryfikacji, a tym samym na efektywność wykorzystania nawozów i ograniczenie strat składników odżywczych.
Jednocześnie intensywna nitryfikacja może przyczyniać się do powstawania gazów cieplarnianych, takich jak podtlenek azotu (N₂O). Dlatego rozwija się praktyki rolnicze mające na celu kontrolę warunków sprzyjających nadmiernej aktywności tych mikroorganizmów, np. poprzez stosowanie inhibitorów nitryfikacji lub odpowiedniego płodozmianu.
Perspektywy przyszłych zastosowań
Wraz z rozwojem biologii syntetycznej pojawia się możliwość modyfikowania szlaków metabolicznych chemoautotrofów w celu tworzenia nowych procesów przemysłowych. Rozważa się np. wykorzystanie ich zdolności do wiązania CO₂ w technologiach sekwestracji węgla lub produkcji biopaliw i bioplastików z wykorzystaniem nieorganicznych donorów energii.
Ciekawym kierunkiem jest też projektowanie bioreaktorów inspirowanych ekosystemami kominów hydrotermalnych, w których organizmy chemoautotroficzne przetwarzałyby odpadowe gazy i ścieki przemysłowe na wartościowe produkty. Prace te znajdują się jednak głównie na etapie badań podstawowych, wymagających dokładniejszego poznania regulacji ich metabolizmu i interakcji z innymi mikroorganizmami.
Znaczenie poznawcze i edukacyjne chemoautotrofów
Zrozumienie chemoautotrofów zmienia perspektywę postrzegania życia. Pokazuje, że biosfera nie jest jedynie cienką, nasłonecznioną warstwą na powierzchni Ziemi, ale obejmuje także głębokie strefy skorupy ziemskiej i dna oceanów. Życie okazuje się zdolne do wykorzystywania różnorodnych źródeł energii, a fotosynteza jest tylko jednym z wielu możliwych rozwiązań.
W edukacji biologicznej wprowadzenie pojęcia chemoautotrofii pomaga lepiej zrozumieć pojęcia takie jak łańcuchy troficzne, cykle biogeochemiczne czy różnorodność metaboliczna mikroorganizmów. Uświadamia też, że mikroorganizmy, choć niewidoczne gołym okiem, mają ogromny wpływ na funkcjonowanie całych ekosystemów i na warunki życia człowieka.
Chemoautotrofy są również doskonałym przykładem związków między biologią a geologią, chemią i fizyką. Ich metabolizm kształtuje minerały i skały, wpływa na skład atmosfery i hydrosfery oraz na przepływy energii w skali globalnej. Interdyscyplinarne badania tych organizmów stanowią ważny element nowoczesnych nauk o Ziemi i życiu.
FAQ – najczęściej zadawane pytania o chemoautotrofy
Czym dokładnie różnią się chemoautotrofy od fotoautotrofów?
Chemoautotrofy i fotoautotrofy łączy zdolność do autotrofii, czyli wytwarzania związków organicznych z nieorganicznego CO₂. Różnią się natomiast źródłem energii. Fotoautotrofy, takie jak rośliny czy sinice, wykorzystują energię promieniowania słonecznego, podczas gdy chemoautotrofy pozyskują ją z reakcji chemicznych utleniania nieorganicznych związków, np. siarkowodoru, jonów amonowych czy żelaza(II). Dzięki temu mogą żyć w ciemności.
Gdzie można znaleźć chemoautotrofy w środowisku naturalnym?
Chemoautotrofy występują przede wszystkim tam, gdzie dostępne są zredukowane związki nieorganiczne i przynajmniej minimalna ilość akceptora elektronów, często tlenu. Spotkać je można w glebie, osadach dennych jezior i mórz, w wodach gruntowych oraz w pobliżu źródeł hydrotermalnych i geotermalnych. Szczególnie liczne są w głębinach oceanicznych przy kominach hydrotermalnych oraz w skałach i wodach podziemnych, gdzie wykorzystują energię geochemiczną.
Jaką rolę pełnią chemoautotrofy w obiegu pierwiastków?
Chemoautotrofy biorą udział w kluczowych etapach obiegu azotu, siarki, żelaza, manganu i węgla. Bakterie nitryfikacyjne przekształcają amon w azotany, które mogą być pobierane przez rośliny. Bakterie siarkowe utleniają siarkowodór do siarczanów, ograniczając jego toksyczność. Mikroorganizmy żelazowe wpływają na rozpuszczalność i osadzanie żelaza w wodach. Dzięki asymilacji CO₂ przyczyniają się też do magazynowania węgla w biomase i osadach, modyfikując skład chemiczny środowiska.
Dlaczego chemoautotrofy są ważne dla teorii powstania życia?
Wiele hipotez zakłada, że pierwsze systemy żywe były organizmami chemolitoautotroficznymi, korzystającymi z energii reakcji zachodzących na powierzchni minerałów w pobliżu kominów hydrotermalnych. Takie środowiska dostarczały stabilnych gradientów chemicznych, które mogły napędzać powstawanie prostych szlaków metabolicznych. Współczesne chemoautotrofy, korzystające z wodoru, siarkowodoru czy CO₂, są traktowane jako modele dawnych form życia i pomagają testować scenariusze prebiotycznej chemii.
Czy chemoautotrofy mogą istnieć poza Ziemią?
Teoretycznie tak, ponieważ do ich funkcjonowania nie jest konieczne światło, a jedynie odpowiednie gradienty chemiczne. Na ciałach niebieskich z podlodowymi oceanami i aktywną geologią, takich jak Europa czy Enceladus, mogą zachodzić reakcje wytwarzające wodór, metan i inne zredukowane związki. Gdyby w takich środowiskach istniały organizmy zdolne do chemoautotrofii, mogłyby tworzyć ekosystemy podobne do tych znanych z kominów hydrotermalnych na dnie Ziemskich oceanów.

