Filamenty pośrednie należą do podstawowych elementów cytoszkieletu komórki eukariotycznej. Tworzą elastyczną, ale wytrzymałą sieć włókien, która zapewnia komórkom odporność mechaniczną, pomaga utrzymać ich kształt oraz organizuje struktury wewnętrzne, takie jak jądro komórkowe. W przeciwieństwie do innych składników cytoszkieletu, filamenty pośrednie nie biorą głównego udziału w ruchu komórki, lecz pełnią funkcję swoistego „rusztowania” i systemu kotwiczącego, kluczowego dla prawidłowego funkcjonowania tkanek.
Budowa i właściwości filamentów pośrednich
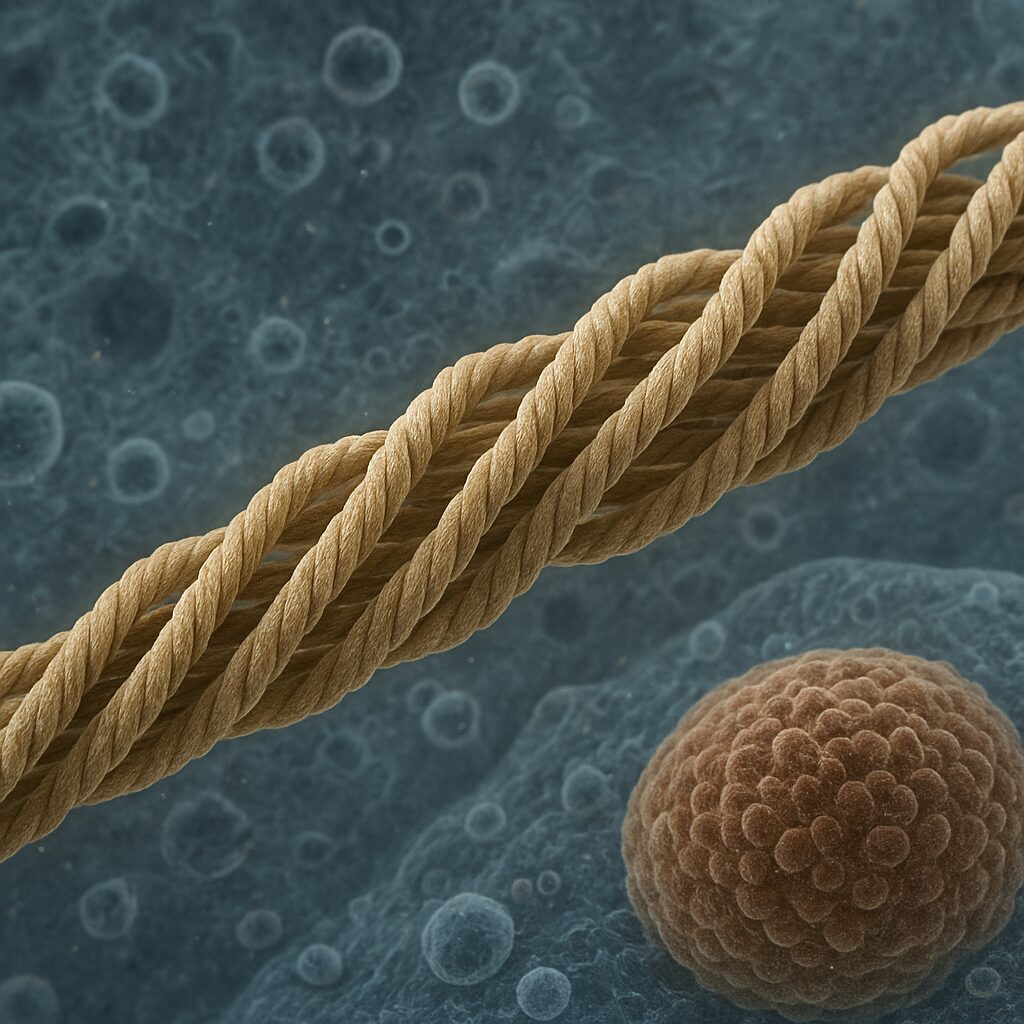
Filamenty pośrednie to wydłużone, włókniste białka o średnicy około 10 nm, co plasuje je pomiędzy cienkimi mikrofilamentami aktynowymi (7 nm) a grubszymi mikrotubulami (25 nm). Ta wielkość stanowi podstawę ich nazwy. Mimo pozornie prostej struktury, filaments pośrednie charakteryzuje złożony sposób organizacji oraz duża różnorodność typów białek, z których są zbudowane. Ich architektura warunkuje unikatowe cechy: wysoką odporność na rozciąganie i ściskanie oraz zdolność do stabilizowania całych tkanek poddawanych ciągłym obciążeniom mechanicznym.
Podstawową jednostką budulcową jest monomer białkowy o wydłużonym kształcie, zawierający charakterystyczną, silnie konserwowaną domenę środkową o strukturze helisy α, otoczoną bardziej zróżnicowanymi końcami N- i C-terminalnymi. Domena helikalna umożliwia powstawanie dimerów – dwóch skręconych wokół siebie cząsteczek. Następnie dimery łączą się w tetramery, a te z kolei układają się bocznie, tworząc protofilamenty. Ostateczna struktura filamentu pośredniego powstaje poprzez boczne i podłużne zestawianie wielu protofilamentów w stabilny, liny przypominający kabel.
Istotną cechą filamentów pośrednich jest ich polimeryzacja, która przebiega inaczej niż w przypadku aktyny czy tubuliny. Nie opiera się ona na zależności od nukleotydów (ATP lub GTP), lecz jest w dużej mierze procesem samorzutnym, regulowanym głównie przez warunki fizykochemiczne i modyfikacje potranslacyjne białek. Dzięki temu włókna te są zazwyczaj trwałe i mniej dynamiczne, co czyni je doskonałymi elementami podporowymi.
Właściwości mechaniczne filamentów pośrednich wynikają zarówno z ich budowy, jak i z liczby oraz rodzaju oddziaływań między poszczególnymi podjednostkami. Do najważniejszych należą:
- wysoka elastyczność – mogą być rozciągane wielokrotnie bez trwałego uszkodzenia struktury;
- zdolność do pochłaniania energii mechanicznej, co amortyzuje działanie sił ścinających i rozciągających;
- duża stabilność – w warunkach in vitro często trudniej je zdemontować niż mikrofilamenty i mikrotubule.
Na końcach nie wykazują one wyraźnej biegunowości, w przeciwieństwie do innych komponentów cytoszkieletu. Brak biegunowości oznacza, że nie stanowią one bezpośredniego toru dla większości białek motorycznych, takich jak kinezyny czy dyneiny, choć mogą wpływać na organizację całej sieci cytoszkieletowej pośrednio, stabilizując położenie innych struktur.
Różnorodność typów i rozmieszczenie filamentów pośrednich
Filamenty pośrednie obejmują wiele rodzin białek, których ekspresja jest silnie zależna od typu komórki, stadium rozwoju oraz stanu fizjologicznego organizmu. Każda grupa tkanek posiada charakterystyczny zestaw białek filamentów pośrednich, czyniąc je nie tylko strukturą mechaniczną, ale również użytecznym markerem różnicowania komórkowego w badaniach biologicznych i diagnostyce patologii, zwłaszcza nowotworów.
Najważniejsze klasy białek filamentów pośrednich to:
- Cytokeratyny (keratyny typu I i II) – występują głównie w komórkach nabłonkowych. Tworzą gęstą sieć łączącą desmosomy oraz hemidesmosomy, co umożliwia przenoszenie sił między komórkami i pomiędzy komórką a macierzą pozakomórkową.
- Wimentyna – charakterystyczna dla komórek mezenchymalnych, takich jak fibroblasty, komórki śródbłonka czy niektóre komórki układu odpornościowego. Ułatwia reorganizację cytoplazmy i uczestniczy w migracji komórek.
- Desmina – obecna w komórkach mięśni szkieletowych, serca i mięśni gładkich. Łączy miofibryle oraz zakotwicza je w błonie komórkowej, zapewniając prawidłowe przenoszenie siły skurczu.
- Neurofilamenty – specyficzne dla neuronów, szczególnie obfite w aksonach, gdzie regulują ich średnicę i przewodnictwo impulsów nerwowych.
- Laminy jądrowe – tworzą sieć tuż pod wewnętrzną błoną otoczki jądrowej. Odpowiadają za utrzymanie kształtu jądra, organizację chromatyny i kontrolę cyklu komórkowego.
Cytokeratyny występują w parach (jedna typu kwaśnego, druga zasadowego), co daje liczne kombinacje heterodimerów. Komórki nabłonkowe różnych narządów wytwarzają odmienne zestawy keratyn, co jest wykorzystywane w histopatologii do określania pochodzenia guza nowotworowego. Na przykład ekspresja określonych keratyn może wskazywać, czy dane komórki wywodzą się z nabłonka przewodu pokarmowego, oddechowego czy skóry.
Wimentyna jest białkiem filamentu pośredniego typowo związanym z komórkami o wysokich zdolnościach migracyjnych i plastyczności fenotypowej. Zwiększona ekspresja wimentyny obserwowana jest podczas przejścia nabłonkowo-mezenchymalnego (EMT), ważnego zarówno w procesach rozwojowych, jak i w progresji nowotworów oraz tworzeniu przerzutów. Jej obecność wiąże się z reorganizacją cytoszkieletu i zmianą kształtu komórki.
Desmina z kolei tworzy sieć otaczającą miofibryle i łączącą je z błoną komórkową, strukturami podporowymi oraz innymi elementami cytoszkieletu. Dzięki temu skurcz jednej miofibryli jest właściwie przekazywany na całą komórkę mięśniową i dalej na całe włókno mięśniowe. Zaburzenia w strukturze lub ekspresji desminy prowadzą do miopatii i kardiomiopatii, objawiających się osłabieniem siły skurczu i degeneracją włókien mięśniowych.
Neurofilamenty pełnią szczególnie istotną funkcję w aksonach. Im większa średnica aksonu, tym szybciej przewodzony jest potencjał czynnościowy. Gęste upakowanie neurofilamentów i ich odpowiednie modyfikacje potranslacyjne (np. fosforylacja) decydują o tej średnicy, a tym samym o szybkości przewodzenia sygnału w układzie nerwowym. Uszkodzenia lub nieprawidłowe odkładanie się neurofilamentów jest związane z chorobami neurodegeneracyjnymi, takimi jak stwardnienie zanikowe boczne (ALS).
Laminy jądrowe (A, C, B1, B2) różnią się od cytoplazmatycznych filamentów pośrednich, lecz zachowują podobną zasadę organizacji. Tworzą one warstwę pod otoczką jądrową, stabilizując jej strukturę i oddziałując z białkami błonowymi oraz chromosomami. Białka te biorą udział w regulacji ekspresji genów, organizacji domen chromatyny oraz mechanicznej odpowiedzi jądra na siły działające na komórkę. Mutacje w genach lamin (zwłaszcza LMNA) wywołują laminopatie, obejmujące m.in. dystrofie mięśniowe, kardiomiopatie oraz pewne formy przedwczesnego starzenia.
Funkcje biologiczne i znaczenie kliniczne filamentów pośrednich
Główna rola filamentów pośrednich polega na nadawaniu komórkom i tkankom odporności mechanicznej. Tworzą one sieć łączącą błonę komórkową z organellami, takimi jak jądro, oraz z innymi komórkami poprzez specjalistyczne struktury adhezyjne. Dzięki temu komórki mogą przetrwać rozciąganie, ściskanie i wibracje, na które są narażone w organizmie, na przykład w skórze, mięśniach czy narządach wewnętrznych.
W nabłonkach filamenty keratynowe są wkomponowane w desmosomy oraz hemidesmosomy. Połączenia te pozwalają na przenoszenie sił działających na pojedynczą komórkę na sąsiednie komórki i na macierz pozakomórkową. Mechanizm ten zapobiega rozrywaniu się tkanki. Gdy keratyny lub białka łączące są nieprawidłowe, nabłonek staje się kruchy i podatny na pęknięcia, co obserwuje się w szeregu chorób dziedzicznych skóry.
W mięśniach sieć desminowa zapewnia spójność struktury sarkomerów w całym włóknie mięśniowym. Bez prawidłowego działania desminy siła generowana przez aparat kurczliwy byłaby słabo przekazywana na całe włókno i tkankę, co prowadziłoby do nieskutecznych skurczów. Odpowiednia organizacja desminy umożliwia także prawidłowe ułożenie mitochondriów oraz innych organelli, co jest niezbędne dla utrzymania zapotrzebowania energetycznego mięśni.
W neuronach neurofilamenty, oprócz roli mechanicznej, tworzą strukturalne „szyny” kształtujące akson. Współdziałają z mikrotubulami i mikrofilamentami aktynowymi, koordynując transport aksonalny pęcherzyków synaptycznych, organelli i białek. Uszkodzenie tej struktury może prowadzić do zaburzeń przewodzenia sygnałów oraz degeneracji włókien nerwowych, co z kolei przekłada się na objawy kliniczne, takie jak osłabienie mięśni, zaburzenia czucia czy upośledzenie funkcji poznawczych.
W jądrze komórkowym laminy odpowiadają nie tylko za mechanikę, ale także za regulację funkcji genomu. Oddziaływanie między lamina jądrową a chromosomami wpływa na organizację przestrzenną chromatyny i ułatwia lub utrudnia dostęp czynników transkrypcyjnych do określonych genów. W konsekwencji, zmiany w strukturze lub ilości lamin mogą modulować profil ekspresji genów i brać udział w procesach różnicowania komórek, starzenia oraz transformacji nowotworowej.
Znaczenie kliniczne filamentów pośrednich jest szczególnie widoczne w przypadku schorzeń wynikających z mutacji w genach kodujących ich białka. Do najlepiej poznanych należą:
- epidermolysis bullosa simplex – choroba pęcherzowa skóry spowodowana mutacjami w genach keratyn K5 lub K14. Łatwo dochodzi do pękania naskórka pod wpływem niewielkich urazów;
- miopatie desminowe – zaburzenia mięśni związane z wadliwą desminą, prowadzące do osłabienia i degeneracji włókien mięśniowych;
- laminopatie – szeroka grupa chorób, m.in. dystrofie mięśniowe obręczy kończyn, kardiomiopatie rozstrzeniowe, lipodystrofie oraz progeria Hutchinsona-Gilforda;
- zaburzenia neurodegeneracyjne z odkładaniem nieprawidłowych agregatów neurofilamentów w neuronach.
Oprócz chorób dziedzicznych, filamenty pośrednie uczestniczą w patogenezie nowotworów. Zmiany w ekspresji poszczególnych typów białek odzwierciedlają często przeprogramowanie komórki nowotworowej. Na przykład komórki raka wywodzące się z nabłonka mogą podczas progresji zacząć eksprymować wimentynę, co jest jednym z przejawów wspomnianego przejścia nabłonkowo-mezenchymalnego. Zwiększona ilość wimentyny wiąże się często z większą inwazyjnością i zdolnością do tworzenia przerzutów.
W praktyce diagnostycznej filamenty pośrednie pełnią rolę istotnych markerów immunohistochemicznych. Techniki barwienia z użyciem przeciwciał przeciwko keratynom, wimentynie lub laminom pozwalają ustalić, z jakiego typu komórki wywodzi się dany nowotwór. Na przykład:
- obecność keratyn wskazuje na pochodzenie nabłonkowe (rak);
- wimentyna sugeruje pochodzenie mezenchymalne (mięsaki) bądź przejście komórek w bardziej migracyjny fenotyp;
- laminy mogą odzwierciedlać zmiany w organizacji jądra, typowe dla pewnych nowotworów.
Pod względem biologii rozwojowej filamenty pośrednie odgrywają istotną rolę w różnicowaniu komórek i kształtowaniu tkanek. W trakcie embriogenezy zmienia się zestaw filamentów pośrednich obecny w danej komórce, co odzwierciedla jej pochodzenie oraz etap dojrzewania. Na przykład komórki macierzyste często eksprymują wimentynę, zaś w miarę różnicowania w kierunku nabłonkowym zaczynają wytwarzać typowe dla siebie keratyny.
Regulacja dynamiki filamentów pośrednich odbywa się poprzez liczne modyfikacje potranslacyjne, takie jak fosforylacja, sumoilacja czy ubikwitynacja. Fosforylacja określonych reszt aminokwasowych może powodować depolimeryzację włókien, co ma znaczenie szczególnie w trakcie mitozy, gdy dochodzi do rozmontowania lamin jądrowych i rozpadu otoczki jądrowej. Po zakończeniu podziału laminy są ponownie defosforylowane i tworzą prawidłową strukturę otoczki wokół nowo powstałych jąder potomnych.
W ostatnich latach coraz większą uwagę poświęca się także roli filamentów pośrednich w odpowiedzi komórki na uszkodzenia DNA i stres mechaniczny. Komórki nieustannie odczuwają oddziaływanie sił fizycznych pochodzących z otoczenia. Transdukcja tych sygnałów z poziomu błony komórkowej do jądra odbywa się między innymi za pośrednictwem sieci cytoszkieletu, w której filamenty pośrednie pełnią rolę „kabli” łączących różne przedziały komórki. Zmiany w ich napięciu i organizacji mogą wpływać na aktywację szlaków sygnalizacyjnych, modyfikacje chromatyny oraz decyzje komórki dotyczące podziału, różnicowania czy apoptozy.
Znajomość biologii filamentów pośrednich ma zatem znaczenie nie tylko dla zrozumienia podstawowej fizjologii komórki, ale również dla opracowywania nowych podejść terapeutycznych. Ukierunkowanie na szlaki regulujące ich organizację i stabilność może stać się elementem strategii leczenia niektórych nowotworów, chorób mięśni czy schorzeń neurodegeneracyjnych. Jednocześnie białka filamentów pośrednich pozostają cennymi biomarkerami, wykorzystywanymi w diagnostyce i monitorowaniu przebiegu chorób.
FAQ
Czym różnią się filamenty pośrednie od innych elementów cytoszkieletu?
Filamenty pośrednie są włóknami o średnicy około 10 nm, pośredniej między mikrofilamentami i mikrotubulami. W odróżnieniu od nich nie wykazują wyraźnej biegunowości i nie zależą bezpośrednio od ATP czy GTP podczas polimeryzacji. Ich główną funkcją jest zapewnianie komórkom odporności mechanicznej i stabilności strukturalnej, a nie generowanie ruchu czy organizowanie transportu wewnątrzkomórkowego. Dlatego pełnią rolę „rusztowania” utrzymującego kształt i integralność tkanek poddawanych obciążeniom.
Dlaczego filamenty pośrednie są ważne w diagnostyce nowotworów?
Każdy typ tkanki charakteryzuje się określonym zestawem białek filamentów pośrednich, np. keratyny w nabłonkach czy wimentyna w tkankach mezenchymalnych. W badaniach histopatologicznych wykorzystuje się przeciwciała rozpoznające te białka, aby określić, z jakiego typu komórek wywodzi się guz. Zmiana profilu ekspresji, np. pojawienie się wimentyny w komórkach pierwotnie nabłonkowych, może wskazywać na większą agresywność nowotworu i zdolność do tworzenia przerzutów, co ma znaczenie prognostyczne i terapeutyczne.
Jakie choroby są związane z mutacjami w białkach filamentów pośrednich?
Mutacje w genach kodujących filamenty pośrednie prowadzą do szeregu chorób zwanych cytoskeletalnymi. Przykładem jest epidermolysis bullosa simplex, w której defekt keratyn powoduje łamliwość naskórka i powstawanie pęcherzy. Zmiany w desminie wywołują miopatie i kardiomiopatie, objawiające się osłabieniem mięśni i niewydolnością serca. Mutacje w genach lamin leżą u podłoża laminopatii, obejmujących dystrofie mięśniowe, lipodystrofie oraz zespoły przedwczesnego starzenia, takie jak progeria Hutchinsona-Gilforda.
Jaką funkcję pełnią laminy w jądrze komórkowym?
Laminy tworzą sieć włókien tuż pod wewnętrzną błoną otoczki jądrowej, stabilizując jej kształt i wytrzymałość. Oddziałują z białkami błonowymi i chromatyną, co wpływa na rozmieszczenie chromosomów oraz dostęp do genów. Podczas mitozy laminy ulegają fosforylacji i depolimeryzacji, co umożliwia rozpad otoczki jądrowej, a następnie jej odtworzenie po podziale. Zmiany w budowie lub ilości lamin mogą zaburzać ekspresję genów, prowadząc do chorób mięśni, serca czy zaburzeń rozwojowych związanych z nieprawidłową organizacją jądra.

