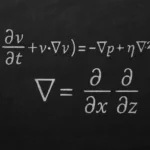
Fotofosforylacja jest jednym z kluczowych procesów biochemicznych na Ziemi, łączącym energię promieniowania świetlnego z syntezą wysokoenergetycznych związków chemicznych. Bez niej nie mogłyby istnieć ani rośliny, ani większość organizmów heterotroficznych, w tym człowiek. Zrozumienie mechanizmów fotofosforylacji pozwala wniknąć w istotę fotosyntezy, a także lepiej pojąć globalny obieg węgla, funkcjonowanie ekosystemów oraz możliwości biotechnologicznego wykorzystania światła słonecznego do produkcji energii i biomasy.
Podstawy fotofosforylacji i jej znaczenie w biologii
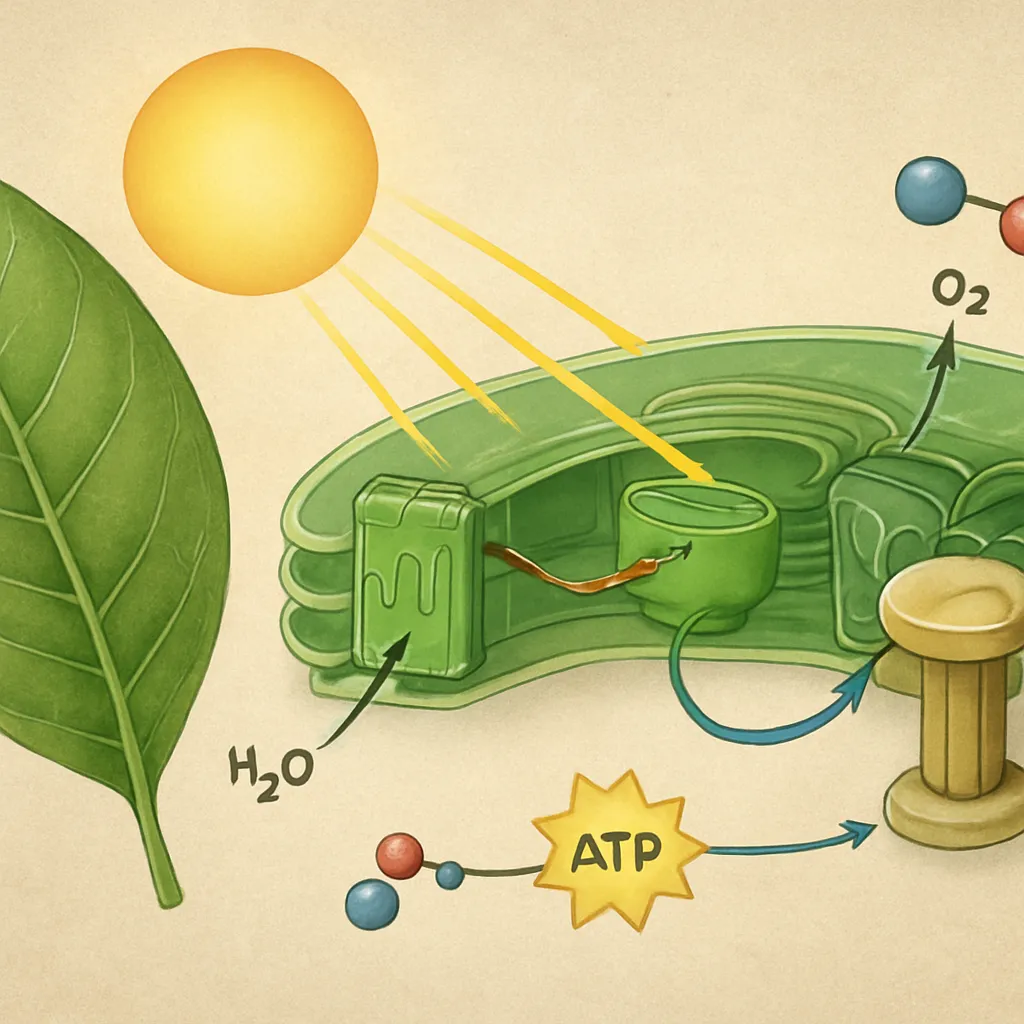
Fotofosforylacja to proces wytwarzania ATP z ADP i nieorganicznego fosforanu (Pi) przy udziale energii światła. Zachodzi w błonach tylakoidów chloroplastów u roślin i glonów, a także w błonach komórkowych bakterii fotosyntetyzujących. Kluczowym elementem tego procesu jest wykorzystanie energii fotonów do napędzania przepływu elektronów i tworzenia gradientu protonowego, który następnie zostaje wykorzystany przez syntazę ATP.
W klasycznym ujęciu fotofosforylacja jest częścią tzw. fazy jasnej fotosyntezy. Podczas tej fazy energia świetlna jest przekształcana w energię chemiczną zmagazynowaną w postaci ATP oraz zredukowanego przenośnika elektronów NADPH. Te związki energetyczne stanowią paliwo dla fazy ciemnej, czyli cyklu Calvina-Bensona, w którym zostaje przeprowadzona asymilacja dwutlenku węgla do związków organicznych.
Z ewolucyjnego punktu widzenia fotofosforylacja umożliwiła organizmom fotosyntetyzującym efektywne wykorzystanie światła jako źródła energii. Rozwój złożonych aparatów fotosyntetycznych oraz centrów reakcji pozwolił na stopniowe zwiększanie sprawności przetwarzania energii i powstanie tlenowej fotosyntezy, odpowiedzialnej za ukształtowanie współczesnej atmosfery bogatej w tlen.
Istnieją dwa główne typy fotofosforylacji: cykliczna i niecykliczna. Różnią się one przebiegiem szlaku transportu elektronów, obecnością lub brakiem produkcji NADPH oraz udziałem poszczególnych fotosystemów. Zależnie od warunków środowiska oraz zapotrzebowania na ATP i NADPH, komórki mogą regulować udział obu typów, aby utrzymać równowagę energetyczną.
Struktury i komponenty odpowiedzialne za fotofosforylację
Podstawową przestrzenią zachodzenia fotofosforylacji u roślin są chloroplasty, a w szczególności ich wewnętrzny system błonowy złożony z tylakoidów. Tylakoidy układają się w granum oraz lamelle stromy, tworząc złożoną sieć, która zapewnia optymalne rozmieszczenie fotosystemów, kompleksów transportu elektronów oraz syntazy ATP. W błonie tylakoidów zakotwiczone są kluczowe białka oraz ich kofaktory, takie jak chlorofile, karotenoidy, feofityny, plastochinon i cytochromy.
Fotosystemy są wyspecjalizowanymi kompleksami białkowo-pigmentowymi, które absorbują światło i inicjują przeniesienie elektronów. U roślin wyższych wyróżnia się fotosystem II (PSII) oraz fotosystem I (PSI). Każdy z nich składa się z centrum reakcji oraz anten barwnikowych. Anteny zawierają liczne cząsteczki chlorofilu a, chlorofilu b oraz karotenoidy, które zbierają energię świetlną i przekazują ją do centrum reakcji w postaci energii wzbudzenia.
Centrum reakcji PSII zawiera specjalną parę chlorofili określaną jako P680, natomiast centrum PSI posiada parę P700. Liczby te odpowiadają długościom fal (w nanometrach), przy których dane fotosystemy wykazują maksymalną absorpcję. Wzbudzenie tych cząsteczek prowadzi do przejścia elektronów na kolejne akceptory, co stanowi początek łańcucha transportu elektronów.
W fotofosforylacji niezwykle ważną rolę odgrywa kompleks cytochromu b6f, pośredniczący między PSII a PSI. To tutaj zachodzi część tzw. cyklu Q, prowadzącego do przepompowywania protonów (H+) przez błonę tylakoidową. Z jednej strony kompleks ten umożliwia transfer elektronów z plastochinolu na plastocyjaninę, z drugiej przyczynia się do budowy gradientu elektrochemicznego protonów, będącego bezpośrednim napędem dla syntazy ATP.
Syntaza ATP jest złożonym enzymem błonowym, który katalizuje fosforylację ADP do ATP. Składa się z dwóch głównych części: F0, będącej kanałem protonowym zanurzonym w błonie, oraz F1, wystającej do strony stromalnej i zawierającej właściwe centra katalityczne. Przepływ protonów przez F0 powoduje obrót podjednostek i mechaniczne wymuszanie cyklu zmian konformacyjnych w części F1, co umożliwia przyłączanie fosforanu do ADP.
Należy podkreślić, że fotofosforylacja, mimo że zachodzi w strukturach wyspecjalizowanych dla fotosyntezy, jest oparta na uniwersalnym mechanizmie chemiosmoticznym opisanym przez Mitchella. Zarówno w chloroplastach, jak i w mitochondriach czy bakteriach, gradient protonowy stanowi źródło potencjału do syntezy ATP. Różnica polega na tym, że w fotosystemach gradient powstaje dzięki energii światła, a w oddychaniu komórkowym – dzięki energii utleniania substratów organicznych.
Mechanizm fotofosforylacji niecyklicznej
Fotofosforylacja niecykliczna, znana także jako nieliniowy transport elektronów, obejmuje współdziałanie fotosystemu II i fotosystemu I. Proces ten prowadzi do równoczesnej syntezy ATP oraz redukcji NADP+ do NADPH, a jego charakterystycznym elementem jest fotoliza wody, związana z uwalnianiem tlenu cząsteczkowego.
Cały cykl można umownie podzielić na kilka etapów. Pierwszym z nich jest absorpcja światła przez anteny barwnikowe PSII. Fotony wzbudzają cząsteczki chlorofilu, a energia wzbudzenia wędruje między pigmentami aż do centrum reakcji P680. Po osiągnięciu centrum wzbudzenie powoduje przejście elektronu na pierwotny akceptor, feofitynę, a następnie na plastochinon (Q).
W wyniku utraty elektronu P680 staje się silnym utleniaczem P680+, zdolnym do odbierania elektronów z kompleksu rozkładającego wodę. Ten tzw. kompleks ewolucji tlenu zawiera mangan i wapń, a jego zadaniem jest kataliza rozszczepienia cząsteczki wody na elektrony, protony oraz tlen. Dwa elektrony przekazywane są z powrotem do P680+, cztery protony gromadzą się w świetle tylakoidu, a jedna cząsteczka O2 uwalniana jest jako produkt uboczny.
Plastochinon, który przyjął elektrony, redukuje się do plastochinolu (PQH2) i dyfunduje w obrębie błony do kompleksu cytochromu b6f. Tam elektrony są kolejno przekazywane przez różne centra redoks, m.in. cytochromy i centrum Fe-S, aż do plastocyjaniny – małego białka miedziowego zlokalizowanego w świetle tylakoidu. W trakcie tych przemian uruchamiany jest cykl Q, powodujący translokację protonów z otoczenia stromalnego do wnętrza tylakoidów, co zwiększa gradient protonowy.
Plastocyjanina dostarcza następnie elektrony do fotosystemu I, gdzie trafiają na centrum reakcji P700. Gdy anteny PSI pochłaniają światło, P700 ulega wzbudzeniu i oddaje elektron do swojego akceptora, którym jest zwykle feofityna lub specjalne centrum chlorofilowe. Następnie elektrony przechodzą przez szereg przenośników, w tym centra żelazowo-siarkowe (Fe-S), aż docierają do białka ferredoksyny.
Ferredoksyna pełni kluczową rolę w końcowej fazie niecyklicznego łańcucha transportu elektronów. Przenosi elektrony na reduktazę ferredoksyna:NADP+, enzym katalizujący reakcję redukcji NADP+ do NADPH. Ten zredukowany koenzym jest niezbędny do późniejszego wiązania CO2 w cyklu Calvina-Bensona. Cały proces niecyklicznego przepływu elektronów tworzy więc ciąg: woda → PSII → cytochrom b6f → PSI → NADP+, z jednoczesną produkcją ATP.
Synteza ATP podczas fotofosforylacji niecyklicznej jest bezpośrednim skutkiem powstawania gradientu protonowego. Protony nagromadzone w świetle tylakoidów, pochodzące zarówno z fotolizy wody, jak i z cyklu Q, tworzą wyższe stężenie H+ wewnątrz niż w stromie. Ta różnica stężeń, uzupełniona o różnicę potencjału elektrycznego po obu stronach błony, określana jest jako siła protonomotoryczna. Przepływ protonów przez kanał F0 syntazy ATP powoduje ciągłe wytwarzanie ATP w części F1.
Istotną zaletą fotofosforylacji niecyklicznej jest równoczesne pozyskiwanie ATP i redukującego NADPH, w proporcjach zbliżonych do zapotrzebowania cyklu Calvina. Jednakże w zależności od warunków środowiskowych, szczególnie intensywności światła oraz dostępności CO2, ta równowaga może zostać zaburzona. W takich sytuacjach znaczenia nabiera fotofosforylacja cykliczna, pozwalająca na dodatkową produkcję ATP bez generacji NADPH.
Mechanizm fotofosforylacji cyklicznej i jej regulacja
Fotofosforylacja cykliczna to alternatywny szlak przepływu elektronów, który angażuje wyłącznie fotosystem I. Elektrony po wzbudzeniu P700 i przeniesieniu na ferredoksynę nie są kierowane do redukcji NADP+, lecz wracają do kompleksu cytochromu b6f lub innych elementów łańcucha, a następnie ponownie do PSI. Taki obieg zamknięty elektronów nie prowadzi do powstawania NADPH ani do fotolizy wody, ale efektywnie generuje gradient protonowy, wykorzystywany przez syntazę ATP do produkcji dodatkowego ATP.
Mechanizm cykliczny pełni ważną funkcję regulacyjną w chloroplastach. Cykl Calvina wymaga nieco większej ilości ATP w stosunku do NADPH, niż jest zazwyczaj dostarczane przez klasyczną fotofosforylację niecykliczną. Gdy w stromie maleje stosunek ATP do NADPH, aktywowane są szlaki cykliczne, które zwiększają wydajność syntezy ATP bez dalszego podnoszenia poziomu zredukowanych ekwiwalentów. Dzięki temu zachowana zostaje równowaga energetyczna i redoks, kluczowa dla sprawnego wiązania dwutlenku węgla.
Jednym z głównych szlaków cyklicznych jest tzw. szlak zależny od kompleksu cytochromu b6f. Ferredoksyna przekazuje w nim elektrony na plastochinon poprzez enzym ferredoksyna:plastochinon reduktaza lub funkcjonalnie równoważne białka. Następnie zredukowany plastochinole przechodzi przez kompleks cytochromu b6f, powodując pompowanie protonów do światła tylakoidu. Efektem tego jest zwiększenie gradientu protonowego i intensyfikacja syntezy ATP, podczas gdy nadmiar redukcyjnego NADPH nie powstaje.
Innym sposobem modulacji cyklicznego przepływu elektronów jest udział białek regulatorowych oraz zmian konformacyjnych anten barwnikowych. Chloroplasty potrafią przemieszczać niektóre anteny między PSI a PSII, dostosowując w ten sposób ilość światła pochłanianego przez poszczególne fotosystemy. Jest to mechanizm tzw. stanów przejściowych (state transitions), który wpływa na rozkład energii pomiędzy liniowym a cyklicznym łańcuchem transportu elektronów.
Cykliczna fotofosforylacja jest także istotnym elementem ochrony aparatu fotosyntetycznego przed fotouszkodzeniami. W warunkach silnego oświetlenia, przy ograniczonej możliwości wykorzystania NADPH w cyklu Calvina (np. wskutek niskiej dostępności CO2), liniowy transport elektronów mógłby prowadzić do nadmiernej redukcji łańcucha i tworzenia reaktywnych form tlenu. Przekierowanie części elektronów na szlaki cykliczne pozwala rozładować nadmiar energii, w bezpieczny sposób podnosząc produkcję ATP i jednocześnie stabilizując stan redoks chloroplastu.
Adaptacyjne znaczenie cyklicznej fotofosforylacji jest szczególnie widoczne u organizmów zamieszkujących środowiska o zmiennym natężeniu światła, takich jak rośliny wyższe w strefie klimatu umiarkowanego czy glony planktonowe w słupie wodnym. Umiejętność elastycznego przełączania się między trybem niecyklicznym a cyklicznym stanowi ważny czynnik decydujący o sprawności fotosyntezy, szybkości wzrostu oraz tolerancji stresu świetlnego.
Zależność fotofosforylacji od czynników środowiskowych
Sprawność fotofosforylacji jest silnie uzależniona od warunków zewnętrznych, w jakich funkcjonuje organizm. Do najważniejszych czynników należą intensywność i jakość światła, temperatura, dostępność wody, koncentracja dwutlenku węgla oraz obecność zanieczyszczeń i czynników stresowych, takich jak metale ciężkie czy herbicydy. Równowaga między lokalnymi warunkami a możliwościami adaptacyjnymi chloroplastów wyznacza granice efektywności fotosyntezy.
Intensywność światła wpływa bezpośrednio na ilość pochłanianej energii przez fotosystemy. Przy niskim natężeniu fotony są czynnikiem ograniczającym i wzrost intensywności prowadzi do wzrostu szybkości fotofosforylacji. Po osiągnięciu wartości nasycającej kolejne zwiększanie ilości światła nie skutkuje wzrostem produkcji ATP, ponieważ ograniczeniem staje się wówczas dostępność innych substratów, głównie CO2, lub pojemność szlaków metabolicznych. Przy bardzo wysokim natężeniu światła organizm musi uruchamiać mechanizmy ochronne, m.in. fotofosforylację cykliczną i niefotochemiczną ekstynkcję, aby uniknąć uszkodzeń centrum reakcji.
Jakość światła, czyli rozkład długości fal, również ma znaczenie. Chlorofil a i b absorbują najbardziej efektywnie w zakresie światła niebieskiego i czerwonego, natomiast słabiej w zakresie zielonym. Co za tym idzie, światło o odpowiednim widmie może zwiększać wydajność pobierania energii i optymalizować proporcje między wzbudzeniem PSII i PSI. W naturalnych warunkach filtracja światła przez liście, wodę czy zawiesiny w środowisku wodnym powoduje zmiany widma, na co organizmy reagują modyfikacją składu barwników i organizacją anten.
Temperatura reguluje kinetykę reakcji enzymatycznych uczestniczących w fotofosforylacji i dalszych etapach fotosyntezy. W umiarkowanym zakresie wzrost temperatury przyspiesza reakcje, aż do osiągnięcia optimum charakterystycznego dla danego gatunku. Po jego przekroczeniu dochodzi do denaturacji białek, destabilizacji kompleksów białkowo-pigmentowych oraz spadku efektywności transportu elektronów. Szczególnie wrażliwy jest kompleks rozkładający wodę oraz niektóre elementy syntazy ATP.
Dostępność wody ma bezpośredni wpływ na fotolizę, ponieważ woda jest substratem dla kompleksu ewolucji tlenu w PSII. Susza ogranicza dopływ wody do chloroplastów, a ponadto powoduje zamykanie aparatów szparkowych, co redukuje dopływ CO2 i pośrednio hamuje wykorzystanie NADPH w cyklu Calvina. To z kolei prowadzi do przeładowania systemu elektronami i wymusza uruchomienie szlaków obronnych, w tym nasilenie cyklicznej fotofosforylacji i powstawanie gradientu protonowego zwiększającego niefotochemiczną ekstynkcję energii.
Nie można pominąć wpływu czynników antropogenicznych, takich jak herbicydy, zanieczyszczenia powietrza i metale ciężkie. Wiele herbicydów działa na poziomie PSII, blokując miejsce wiązania plastochinonu i uniemożliwiając transport elektronów w dół łańcucha. Skutkiem tego jest spadek produkcji ATP i NADPH, a w konsekwencji zahamowanie biosyntezy związków organicznych i obumarcie rośliny. Z kolei niektóre metale ciężkie mogą zaburzać skład centrów reakcji, uszkadzać kompleksy białkowe i generować stres oksydacyjny, co prowadzi do degradacji aparatu fotosyntetycznego.
Znaczenie fotofosforylacji w ekologii i biotechnologii
Fotofosforylacja ma fundamentalne znaczenie nie tylko na poziomie biochemicznym, lecz także w skali całej biosfery. Zdolność organizmów fotosyntetyzujących do przekształcania energii promieniowania słonecznego w energię chemiczną warunkuje funkcjonowanie większości ekosystemów. Rośliny, glony i cyjanobakterie stanowią podstawę piramid troficznych, dostarczając energii i materii organicznej wyższym poziomom, takim jak roślinożercy czy drapieżniki.
Przemiany związane z fotofosforylacją ściśle wiążą się z globalnym cyklem węgla. Produkcja ATP i NADPH w fazie jasnej jest konieczna do redukcji CO2 w cyklu Calvina i tworzenia węglowodanów. W długich skalach czasowych akumulacja biomasy roślinnej oraz jej częściowe przekształcenie w paliwa kopalne zadecydowały o obecnym bilansie węgla w atmosferze, hydrosferze i litosferze. Zrozumienie mechanizmów kontroli fotofosforylacji pomaga przewidywać reakcje ekosystemów na zmiany klimatu oraz potencjalne sprzężenia zwrotne między biosferą a atmosferą.
W biotechnologii fotofosforylacja jest obiektem intensywnych badań z kilku powodów. Po pierwsze, rośnie zainteresowanie wykorzystaniem roślin i mikroalg jako systemów produkcyjnych do biopaliw, bioplastików i innych związków o znaczeniu przemysłowym. Zwiększenie wydajności konwersji światła w ATP oraz węgla w biomasę może przełożyć się na lepszą opłacalność takich technologii. Inżynieria genetyczna chloroplastów i modyfikowanie elementów łańcucha transportu elektronów to jedna z dróg optymalizacji.
Po drugie, badania nad sztuczną fotosyntezą inspirują się naturalnymi mechanizmami fotofosforylacji. Celem jest stworzenie układów, które podobnie jak fotosystemy potrafią stabilnie i wydajnie przetwarzać energię światła na użyteczne formy chemiczne, np. wodór, alkohole czy inne paliwa. W tym kontekście mechanizmy tworzenia gradientu protonowego i katalityczna funkcja syntazy ATP stanowią ważny punkt odniesienia przy projektowaniu biomimetycznych ogniw słonecznych i reaktorów fotochemicznych.
Po trzecie, zrozumienie regulacji fotofosforylacji ma znaczenie w rolnictwie i hodowli roślin. Możliwość sterowania równowagą między fotofosforylacją cykliczną a niecykliczną, a także zwiększanie odporności aparatu fotosyntetycznego na stres świetlny, suszę czy skrajne temperatury, może przyczynić się do tworzenia odmian o wyższej wydajności plonowania i większej stabilności w zmieniającym się klimacie.
Wreszcie, fotofosforylacja jest też obiektem badań podstawowych w zakresie bioenergetyki. Pozwala testować ogólne hipotezy dotyczące sprzęgania energii, dynamiki białek błonowych, roli wody w procesach biologicznych i powstawania struktur o złożonej organizacji. Bioenergetyka fotosyntetyczna dostarcza wielu modeli teoretycznych i danych eksperymentalnych, które są ważne również dla zrozumienia funkcjonowania mitochondriów, bakterii chemosyntetyzujących oraz innych systemów wykorzystujących gradienty jonowe.
Różnorodność fotofosforylacji u różnych grup organizmów
Choć ogólny mechanizm fotofosforylacji jest wspólny dla wielu organizmów, szczegóły strukturalne i funkcjonalne mogą się istotnie różnić. U roślin wyższych aparat fotosyntetyczny znajduje się w chloroplastach, które wyewoluowały z endosymbiotycznych cyjanobakterii. Współczesne cyjanobakterie posiadają podobny zestaw fotosystemów i łańcuchów transportu elektronów jak rośliny, lecz ich fotofosforylacja zachodzi w złożonych systemach błonowych wewnątrz cytoplazmy, zwanych tylakoidami bakteryjnymi.
Inne bakterie fotosyntetyzujące, takie jak bakterie purpurowe czy zielone bakterie siarkowe, korzystają z odmiennych barwników (bakteriochlorofile) oraz często nie posiadają tlenowej fotolizy wody. Zamiast tego wykorzystują alternatywne donory elektronów, np. siarkowodór, i prowadzą beztlenową fotosyntezę. Ich fotofosforylacja opiera się najczęściej na cyklicznym transporcie elektronów z udziałem pojedynczego fotosystemu, a gradient protonowy powstaje dzięki przepompowywaniu protonów i transporcie elektronów wewnątrz jednego kompleksu.
Glony jednokomórkowe, takie jak Chlamydomonas reinhardtii, stanowią interesujący model badań, ponieważ łączą cechy roślin lądowych i prostych eukariontów. Ich zdolność do szybkiej zmiany trybu metabolicznego, przechodzenia między fotosyntezą a oddychaniem heterotroficznym oraz wysoka plastyczność fotosystemów sprawiają, że są często wykorzystywane w badaniach nad adaptacją fotofosforylacji do różnych warunków oświetlenia i stężenia tlenu.
Wiele organizmów wykazuje dodatkowe formy fotofosforylacji adaptacyjnej, np. związanej z fototropizmem, fotomorfogenezą czy mechanizmami zegara biologicznego. U roślin wyższych działanie fotoreceptorów niebieskiego i czerwonego światła (kryptochromy, fototropiny, fitohromy) współgra z funkcją fotosystemów, kształtując architekturę liści, orientację aparatów szparkowych i rozmieszczenie chloroplastów w komórce. Wszystkie te mechanizmy w sposób pośredni wpływają na efektywność transportu elektronów i stopień wykorzystania gradientu protonowego.
Nawet u organizmów nieposiadających klasycznej fotosyntezy mogą występować elementy systemów fotochemicznych, takie jak rodopsyny bakteryjne, które pompują protony pod wpływem światła i umożliwiają syntezę ATP. Choć nie są one częścią pełnego aparatu fotosyntetycznego, ich działanie opiera się na podobnej zasadzie chemiosmotycznej. Stanowią przykład konwergentnej ewolucji mechanizmów wykorzystania energii promieniowania do budowy gradientu jonowego i napędu dla enzymów syntetyzujących ATP.
FAQ – najczęściej zadawane pytania o fotofosforylację
Czym różni się fotofosforylacja od fosforylacji oksydacyjnej?
Fotofosforylacja zachodzi w chloroplastach i polega na wykorzystaniu energii światła do pompowania protonów i syntezy ATP. Fosforylacja oksydacyjna odbywa się w mitochondriach i jest napędzana energią z utleniania związków organicznych. W pierwszym przypadku źródłem elektronów jest woda, a głównym produktem obok ATP jest NADPH; w drugim – zredukowane koenzymy (NADH, FADH2), a produktem końcowym jest woda i ATP.
Dlaczego fotofosforylacja jest tak ważna dla życia na Ziemi?
Fotofosforylacja dostarcza ATP i siły redukcyjnej potrzebnej do wiązania CO2 w złożone związki organiczne. Dzięki temu rośliny, glony i cyjanobakterie tworzą biomasę stanowiącą pokarm dla pozostałych organizmów. Proces ten uwalnia również tlen do atmosfery, umożliwiając rozwój oddychania tlenowego. Bez fotofosforylacji nie istniałby obecny łańcuch troficzny, a większość ekosystemów uległaby załamaniu.
Czym jest gradient protonowy i jak napędza syntezę ATP?
Gradient protonowy to różnica stężenia protonów (H+) i potencjału elektrycznego po obu stronach błony tylakoidowej. Podczas transportu elektronów protony są pompowane do światła tylakoidów, tworząc nadwyżkę ładunku dodatniego wewnątrz. Dążąc do wyrównania gradientu, protony wracają do stromy przez kanał syntazy ATP. Energia ich przepływu zostaje przekształcona w energię wiązań chemicznych ATP, co opisuje teoria chemiosmotyczna.
Dlaczego istnieje fotofosforylacja cykliczna i niecykliczna?
Oba typy fotofosforylacji pełnią uzupełniające się funkcje. Niecykliczna zapewnia jednocześnie ATP, NADPH i tlen, zaspokajając podstawowe potrzeby fotosyntezy. Cykliczna służy głównie korygowaniu bilansu energetycznego: pozwala produkować dodatkowe ATP bez tworzenia NADPH, co jest konieczne, gdy cykl Calvina wymaga więcej energii fosforanowej niż redukcyjnej. Dodatkowo cykliczny przepływ elektronów pomaga chronić aparat fotosyntetyczny przed fotouszkodzeniami.
Jak człowiek może wykorzystać wiedzę o fotofosforylacji?
Wiedza o fotofosforylacji znajduje zastosowanie w rolnictwie, biotechnologii i energetyce. Pozwala tworzyć rośliny bardziej odporne na stres i wydajniejsze w produkcji biomasy, optymalizować uprawy szklarniowe pod względem oświetlenia, a także projektować mikroalgi jako bioreaktory do wytwarzania biopaliw czy farmaceutyków. Inspiruje też konstrukcję sztucznych systemów fotosyntetycznych, zdolnych do konwersji światła słonecznego w paliwa chemiczne na wzór naturalnych fotosystemów.