Kataliza enzymatyczna stanowi jeden z najbardziej niezwykłych przejawów organizacji materii w świecie przyrody. Enzymy, będące wyspecjalizowanymi biokatalizatorami, umożliwiają zachodzenie reakcji chemicznych w warunkach, w których bez ich udziału przebiegałyby ekstremalnie wolno lub wcale. Dzięki nim komórki mogą prowadzić złożone przemiany energetyczne i budulcowe, utrzymywać homeostazę oraz reagować na bodźce środowiskowe z ogromną precyzją. Zrozumienie natury katalizy enzymatycznej łączy wiedzę z zakresu chemii fizycznej, biochemii, biologii molekularnej oraz inżynierii procesowej, tworząc fundament nowoczesnych nauk przyrodniczych i technologii biotechnologicznych.
Podstawy katalizy enzymatycznej – od energii aktywacji do stanu przejściowego
Każda reakcja chemiczna, aby mogła zajść, musi pokonać barierę energetyczną zwaną energią aktywacji. Nawet jeśli różnica energii między substratami a produktami wskazuje, że proces jest termodynamicznie korzystny, to bez odpowiedniego obniżenia bariery, liczba cząsteczek osiągających stan przejściowy pozostaje niewielka. W układach nieożywionych rolę katalizatorów pełnią metale, kwasy mineralne czy kompleksy koordynacyjne. W organizmach żywych funkcję tę przejęły wyspecjalizowane białka – enzymy.
Enzym nie zmienia całkowitej energii swobodnej reakcji (ΔG), lecz modyfikuje drogę reakcji, wprowadzając alternatywny mechanizm prowadzący przez niżej położony stan przejściowy. Oznacza to, że katalizator enzymatyczny nie „wypycha” układu poza prawa termodynamiki, lecz umożliwia wykorzystanie korzystnych przemian w czasie istotnym biologicznie. Z perspektywy chemii fizycznej istotne jest, że obniżenie energii aktywacji skutkuje wzrostem stałej szybkości reakcji zgodnie z równaniem Arrheniusa, a zatem drastyczną zmianą kinetyki przy niezmienionej równowadze termodynamicznej.
Szczególnie ważne jest pojęcie stanu przejściowego. W teorii katalizy enzymatycznej przyjmuje się, że enzym najsilniej stabilizuje nie tyle substrat czy produkt, ile właśnie stan przejściowy – układ o częściowo rozerwanych i częściowo utworzonych wiązaniach. Dość proste rozumowanie pokazuje, że jeżeli białko zostało tak zorganizowane, aby jego centrum aktywne komplementarnie wiązało strukturę przypominającą stan przejściowy, to energia potrzebna do jego osiągnięcia spada. Stąd wynika jedna z kluczowych tez: enzymy to katalizatory stanu przejściowego.
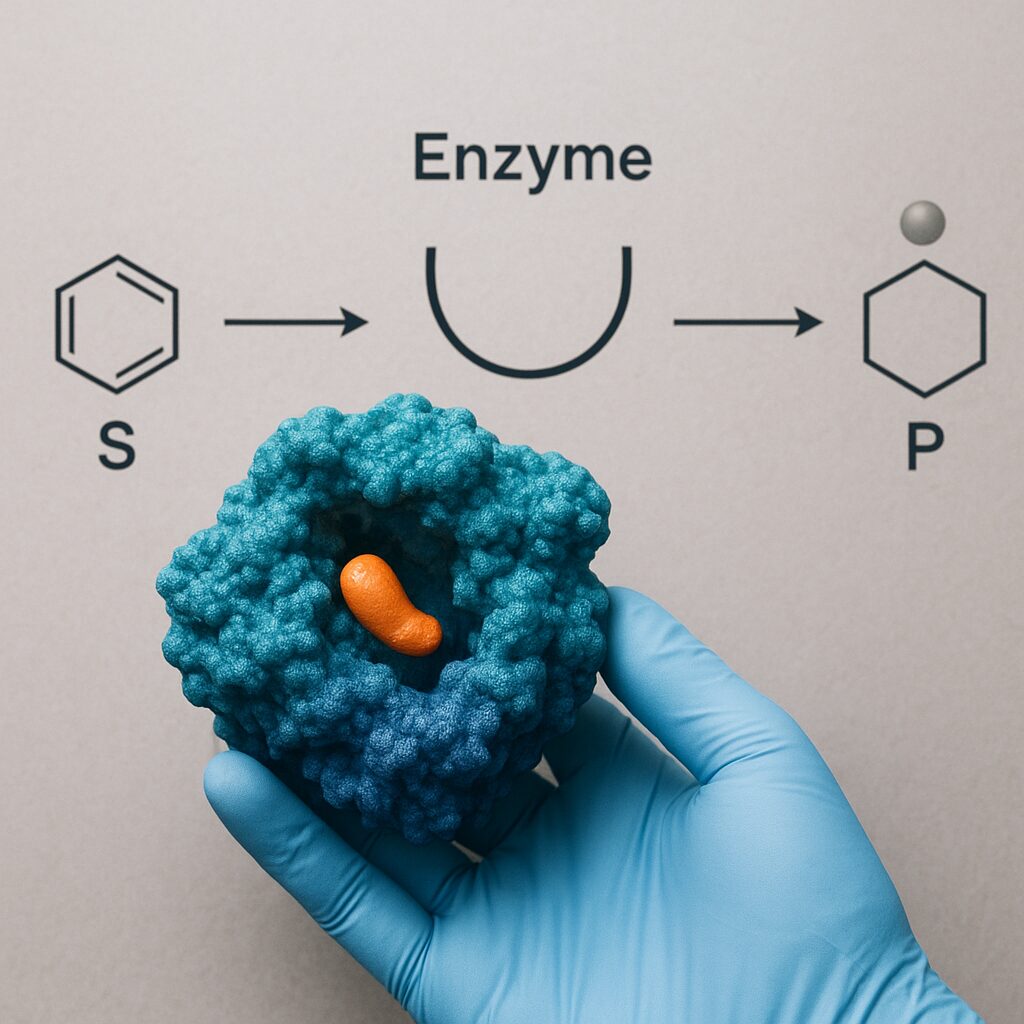
Z punktu widzenia chemii strukturalnej enzymy są zwykle zbudowane z jednego lub kilku łańcuchów polipeptydowych o określonej strukturze drugorzędowej, trzeciorzędowej i niekiedy czwartorzędowej. Ułożenie helis α, harmonijek β i pętli decyduje o powstaniu zagłębień i kieszeni, spośród których najważniejsza jest kieszeń katalityczna – centrum aktywne. To tam dochodzi do wiązania substratu i przeprowadzenia reakcji chemicznej.
Należy podkreślić, że kataliza enzymatyczna nie sprowadza się do prostego „przechwycenia” reagenta. Enzymy wykorzystują szereg zjawisk fizykochemicznych: specyficzne oddziaływania elektrostatyczne, indukowanie napięcia w wiązaniach, koordynację z metalami przejściowymi, przeniesienie protonów czy udział nukleofilowych grup bocznych aminokwasów. Wszystko to odbywa się przy temperaturze i ciśnieniu typowym dla organizmów, najczęściej w roztworach wodnych, które same w sobie silnie wpływają na przebieg reakcji.
Klasyczna analiza kinetyki enzymatycznej opiera się na modelu Michaelisa-Menten, który wprowadza dwa podstawowe parametry: stałą Michaelisa (Km) oraz maksymalną szybkość reakcji (Vmax). Choć jest to ujęcie uproszczone, zakładające m.in. istnienie stanu quasi-równowagi między enzymem, substratem i kompleksem pośrednim, pozwala ono na ilościowy opis szybkości przekształceń przy różnych stężeniach substratu oraz na porównywanie aktywności różnych enzymów lub ich wariantów.
Specyficzność enzymów i modele dopasowania w centrum aktywnym
Unikalną cechą katalizy enzymatycznej jest wysoka specyficzność działania. O ile katalizatory nieorganiczne przyspieszają zwykle całe klasy reakcji, o tyle pojedynczy enzym może być wyspecjalizowany w przemianie jednego typu wiązania w bardzo określonym substracie. Ta wyjątkowość wynika z komplementarności przestrzennej i chemicznej między centrum aktywnym a reagującą cząsteczką.
Historycznie rozwój teorii specyficzności zaczął się od modelu „klucza i zamka” zaproponowanego przez Emila Fischera. W tym prostym ujęciu kształt centrum aktywnego jest idealnie dopasowany do kształtu substratu, podobnie jak zamek jest zaprojektowany pod konkretny klucz. Model dobrze tłumaczy wysoką selektywność, ale jest zbyt sztywny, aby oddać rzeczywistość dynamicznych makrocząsteczek białkowych.
Nowocześniejsze podejście określane mianem „dopasowania indukowanego” (induced fit) zakłada, że po początkowym, wstępnym związaniu substratu, enzym ulega zmianom konformacyjnym prowadzącym do optymalizacji kontaktu w centrum aktywnym. Substrat niejako „wymusza” korzystne ustawienie reszt aminokwasowych, które biorą udział w katalizie. Pozwala to wyjaśnić, dlaczego wiele enzymów jest w stanie rozróżniać nawet bardzo podobne cząsteczki, ponieważ tylko jedna z nich indukuje właściwy zestaw zmian strukturalnych.
W centrum aktywnym można wyróżnić dwie strefy: obszar wiążący substrat oraz obszar właściwie katalityczny, w którym znajdują się reszty bezpośrednio uczestniczące w tworzeniu i zrywaniu wiązań chemicznych. Wśród najczęściej zaangażowanych aminokwasów znajdują się histydyna, seryna, cysteina, kwas glutaminowy, kwas asparaginowy oraz lizyna. Ich grupy boczne zachowują się jak silne nukleofile, kwasy Bronsteda lub zasady, umożliwiając protonowanie, deprotonowanie czy atak na centra elektrofilowe substratu.
Specyficzność dotyczy nie tylko rodzaju substratu, lecz także jego konfiguracji stereochemicznej. W środowisku biologicznym szczególne znaczenie ma chiralność – wiele enzymów rozpoznaje wyłącznie izomery L-aminokwasów lub D-cukrów, co przekłada się na jednolity charakter makrocząsteczek w żywych organizmach. Z chemicznego punktu widzenia oznacza to zdolność do niezwykle precyzyjnego rozróżniania przestrzennych ułożeń grup funkcjonalnych w obrębie tej samej formuły sumarycznej.
Warto podkreślić, że centrum aktywne jest często wspierane przez elementy strukturalne położone daleko w sekwencji pierwotnej. Zmiany w odległych fragmentach białka mogą modulować dostępność kieszeni katalitycznej, jej ładunek powierzchniowy lub elastyczność. To tłumaczy, dlaczego nawet pozornie małe mutacje, nienaruszające bezpośrednio reszt katalitycznych, mogą prowadzić do istotnych zmian aktywności, lub nawet całkowitej utraty funkcji enzymu.
Zjawisko to ma ogromne znaczenie w chemii białek i inżynierii enzymów. Projektowanie wariantów o zmienionej specyficzności – na przykład takich, które preferują inny substrat przemysłowy – wymaga nie tylko intuicji dotyczącej miejsca aktywnego, lecz także uwzględnienia całej „architektury” białka. Stąd rosnące znaczenie metod bioinformatycznych, modelowania molekularnego oraz badań strukturalnych (np. krystalografia rentgenowska, NMR, krio-EM) w projektowaniu nowych katalizatorów białkowych.
Klasy dobrego dopasowania enzym-substrat można dodatkowo porządkować według rodzaju reakcji. Hydrolazy rozpoznają wiązania podatne na atak nukleofila wodnego, oksydoreduktazy selektywnie kierują transferem elektronów i protonów, transferazy przenoszą określone grupy funkcyjne, a liazy czy izomerazy wymagają subtelnego ustawienia substratu, aby umożliwić sprzężone rozerwanie i tworzenie wiązań bez udziału klasycznego nukleofila. Każda grupa enzymów reprezentuje inną strategię dopasowania, choć łączy je wspólny cel – stabilizacja struktury zbliżonej do stanu przejściowego.
Mechanizmy działania enzymów – strategie chemiczne i czynniki wpływające na aktywność
Kataliza enzymatyczna opiera się na kilku głównych strategiach chemicznych, które często występują równocześnie. Jednym z najważniejszych jest kataliza kwasowo-zasadowa, w której grupy funkcyjne aminokwasów pełnią rolę donorów lub akceptorów protonów. Histydyna, dzięki pKa swojego pierścienia imidazolowego bliskiemu fizjologicznemu pH, bywa szczególnie efektywnym uczestnikiem tego typu mechanizmów. Umożliwia ona tworzenie przejściowych form protonowanych i zdeprotonowanych, stabilizujących stan przejściowy w reakcji hydrolyzy, izomeryzacji czy kondensacji.
Drugim typem jest kataliza kowalencyjna, polegająca na tworzeniu przejściowego wiązania między enzymem a substratem. Przykładem mogą być proteazy serynowe, takie jak chymotrypsyna czy trypsyna, w których reszta seryny atakuje nukleofilowo wiązanie peptydowe, tworząc krótkotrwały acylenzym. W kolejnym etapie dochodzi do hydrolizy tego kompleksu i odtworzenia wolnego enzymu. Z chemicznego punktu widzenia mechanizm ten przypomina klasyczne reakcje acylowania i deacylowania obserwowane w chemii organicznej.
Kolejna strategia wiąże się z katalizą elektrostatyczną. Enzym wykorzystuje rozmieszczenie ładunków w centrum aktywnym, aby stabilizować naładowane formy przejściowe, jony oksoniowe lub karbokationowe, a także aby kierować substrat do optymalnego położenia. Oddziaływania jonowe, dipolowe i wiązania wodorowe tworzą złożoną sieć, która zmniejsza energię swobodną struktury przejściowej. Wiele badań wskazuje, że właśnie precyzyjne dostrojenie potencjału elektrostatycznego w centrum aktywnym jest jednym z najtrudniejszych do odtworzenia aspektów przy projektowaniu sztucznych katalizatorów.
Warto podkreślić także rolę katalizy metalozależnej. Liczne enzymy zawierają jony metali, takie jak Zn²⁺, Mg²⁺, Fe²⁺/Fe³⁺, Cu²⁺ czy Mn²⁺, koordynowane przez reszty aminokwasowe i dodatkowe ligandy. Metale te pełnią różnorodne funkcje: aktywują cząsteczki wody, ułatwiają przeniesienie ladunku, stabilizują ujemnie naładowane stany przejściowe lub uczestniczą w wieloetapowych reakcjach redoks. Przykładem są dehydrogenazy zależne od cynku czy polimerazy DNA wymagające jonów magnezu do prawidłowej elongacji łańcucha nukleotydowego.
Na aktywność enzymów silnie wpływa środowisko reakcji. Temperatury zbyt wysokie prowadzą do denaturacji białka – zniszczenia jego struktury trzeciorzędowej i utraty funkcjonalnego centrum aktywnego. Zbyt niskie temperatury ograniczają ruchy konformacyjne, spowalniając procesy wiązania substratu i uwalniania produktu. Optimum temperaturowe wielu enzymów ludzkich mieści się w wąskim przedziale około 37°C, co odzwierciedla warunki fizjologiczne organizmu.
Podobnie krytyczne jest pH środowiska. Każdy enzym ma określone optimum pH, w którym jonizacja reszt katalitycznych sprzyja przeprowadzeniu reakcji. Odchylenia od tego optimum skutkują niewłaściwym protonowaniem, zaburzeniem sieci wiązań wodorowych oraz zmianą rozpuszczalności substratu. Przykładowo, pepsyna żołądkowa funkcjonuje najefektywniej w silnie kwaśnym środowisku, podczas gdy enzymy trzustkowe wymagają warunków zbliżonych do obojętnych w jelicie cienkim.
Stężenie substratu stanowi kolejny istotny czynnik. Wzrost stężenia początkowo powoduje niemal proporcjonalny wzrost szybkości reakcji, aż do osiągnięcia stanu wysycenia, kiedy wszystkie centra aktywne są zajęte. W tym momencie szybkość maksymalna (Vmax) jest wprost zależna od liczby aktywnych cząsteczek enzymu, a nie od ilości substratu. Stała Michaelisa (Km) odzwierciedla z kolei powinowactwo enzymu do substratu – im mniejsza jej wartość, tym silniej i efektywniej białko wiąże reagenta.
W naturalnych układach biologicznych aktywność enzymów jest regulowana wieloma mechanizmami. Inhibicja kompetycyjna polega na rywalizacji między substratem a cząsteczką podobną strukturalnie, która jednak nie ulega przemianie. W inhibicji niekompetycyjnej inhibitor wiąże się w miejscu innym niż centrum aktywne, zmieniając konformację enzymu i obniżając jego maksymalną aktywność. Z kolei aktywacja allosteryczna wykorzystuje obecność wyspecjalizowanych miejsc wiążących, których obsadzenie przez ligandy sprzyja przyjęciu formy o wyższej aktywności katalitycznej.
Regulacja ta ma fundamentalne znaczenie dla utrzymania równowagi metabolicznej. Produkty końcowe szlaków biosyntetycznych często działają jako inhibitory allosteryczne enzymów z pierwszych etapów ścieżki, co zapobiega nadmiernej akumulacji zbędnych związków. Tego typu sprzężenie zwrotne ujemne jest klasycznym przykładem subtelnej integracji katalizy enzymatycznej z potrzebami całego organizmu.
Rozumienie mechanizmów katalizy enzymatycznej ma ogromne implikacje praktyczne. Projektowanie leków polega często na opracowaniu cząsteczek będących przejściowymi odpowiednikami stanu przejściowego, które wiążą się z enzymem silniej niż naturalny substrat, hamując jego działanie. Inżynieria białek pozwala natomiast na tworzenie wariantów bardziej odpornych na wysoką temperaturę czy ekstremalne pH, co jest kluczowe dla zastosowań przemysłowych w chemii organicznej, produkcji żywności czy biopaliw.
Zastosowania i perspektywy katalizy enzymatycznej w nauce i technologii
Kataliza enzymatyczna przeniknęła daleko poza klasyczną biochemię komórki, stając się narzędziem chemii stosowanej, biotechnologii oraz inżynierii materiałowej. W świecie przemysłu spożywczego enzymy odpowiadają za klarowanie soków, modyfikację skrobi, zmiękczanie mięsa czy tworzenie odpowiedniej struktury pieczywa. Proteazy, amylazy i lipazy są powszechnie dodawane do detergentów, co pozwala na skuteczne usuwanie plam białkowych i tłuszczowych w niższych temperaturach prania, a tym samym na ograniczenie zużycia energii.
W syntezie organicznej enzymy wykorzystuje się jako wysoce selektywne katalizatory w reakcjach tworzenia wiązań estrowych, amidowych, etherowych czy C–C. Szczególnie cenne są ich właściwości stereoselektywne, umożliwiające otrzymywanie pojedynczych enancjomerów leków. W dobie rosnącej świadomości ekologicznej procesy biokatalityczne zyskują na znaczeniu jako alternatywa dla klasycznej chemii opartej na metalach ciężkich, toksycznych rozpuszczalnikach i wysokich temperaturach. Biokataliza pozwala na projektowanie „zielonych” ścieżek technologicznych, zgodnych z zasadami zrównoważonego rozwoju.
W naukach medycznych kataliza enzymatyczna stanowi bezpośredni cel terapii farmakologicznych. Liczne leki działają jako inhibitory kluczowych enzymów patogenów, np. proteazy wirusowe czy polimerazy kwasów nukleinowych. Precyzyjne poznanie mechanizmów działania enzymu chorobotwórczego umożliwia zaprojektowanie molekuł, które blokują aktywne centrum lub zakłócają niezbędne przejścia konformacyjne. Równocześnie badania nad ludzkimi enzymami ujawniają, w jaki sposób mutacje genetyczne zmieniają parametry kinetyczne i prowadzą do chorób metabolicznych.
Szczególne miejsce zajmuje inżynieria enzymów, czyli celowa modyfikacja sekwencji aminokwasowych w celu uzyskania nowych własności katalitycznych. Wykorzystuje się tu zarówno klasyczne mutagenezy ukierunkowane na określone pozycje, jak i bardziej złożone metody ewolucji kierowanej, w których tworzy się biblioteki zmutowanych wariantów i selekcjonuje te o najkorzystniejszych parametrach. Pozwala to opracowywać enzymy aktywne w rozpuszczalnikach organicznych, stabilne termicznie lub zdolne do przyspieszania reakcji niewystępujących naturalnie.
Na granicy chemii i biologii molekularnej rozwija się też dziedzina ribozymów i deoksyrybozymów – katalitycznych kwasów nukleinowych. Choć klasycznie katalizatorami biologicznymi są białka, okazało się, że odpowiednio ukształtowane RNA i DNA mogą przyspieszać określone reakcje, np. cięcie i ligację wiązań fosfodiestrowych. To otwiera drogę do zrozumienia pierwotnych etapów ewolucji życia, w których kwasy nukleinowe mogły jednocześnie przechowywać informację genetyczną i pełnić funkcje katalityczne, tworząc hipotetyczny „świat RNA”.
Nowoczesna nanotechnologia również sięga po katalizę enzymatyczną. Enzymy immobilizowane na powierzchniach nanomateriałów, takich jak nanorurki węglowe, tlenki metali czy polimery przewodzące, tworzą biosensory umożliwiające wykrywanie śladowych ilości związków chemicznych. Klasycznym przykładem jest glukozoksydaza wykorzystywana w paskach testowych do monitorowania poziomu glukozy we krwi. Zrozumienie zasad działania enzymów pomaga tu projektować układy o optymalnym przeniesieniu elektronów między centrum aktywnym a elektrodą pomiarową.
W obszarze energetyki rosnące zainteresowanie budzą enzymy uczestniczące w przemianach związków bogatoenergetycznych. Dehydrogenazy, oksydazy i hydrogenazy są rozpatrywane jako komponenty ogniw paliwowych opartych na biokatalizie, zdolnych do efektywnego przetwarzania paliw odnawialnych. Badania nad kompleksami enzym-nanomateriał-metal mają na celu pokonanie ograniczeń związanych z trwałością i wydajnością tych systemów, a w dalszej perspektywie stworzenie urządzeń łączących zalety tradycyjnej elektrokatalizy i wysokiej selektywności enzymów.
Kataliza enzymatyczna znajduje też zastosowanie w ochronie środowiska. Enzymy degradujące pestycydy, związki ropopochodne czy barwniki przemysłowe są wykorzystywane w procesach bioremediacji. Dzięki swojej wysokiej selektywności mogą rozkładać zanieczyszczenia do prostszych, mniej szkodliwych produktów, często w warunkach łagodniejszych niż w klasycznych metodach fizykochemicznych. Szczególnie interesujące są systemy wykorzystujące mikroorganizmy inżynierowane genetycznie, wyposażone w zestawy enzymów dostosowanych do specyficznych zanieczyszczeń lokalnych.
Analizując perspektywy badań, można spodziewać się dalszej integracji katalizy enzymatycznej z projektowaniem obwodów metabolicznych na poziomie całych organizmów. Biologia syntetyczna dąży do konstruowania komórek o nowatorskich szlakach biochemicznych, zdolnych do produkcji związków farmaceutycznych, polimerów funkcjonalnych czy biopaliw z prostych substratów, takich jak dwutlenek węgla czy odpadowa biomasa. Osiągnięcie tego celu wymaga precyzyjnego zrozumienia i kontrolowania aktywności wielu enzymów jednocześnie, co czyni katalizę enzymatyczną jednym z centralnych zagadnień współczesnej chemii i nauk pokrewnych.
FAQ – najczęstsze pytania o katalizę enzymatyczną
Na czym polega różnica między katalizą enzymatyczną a nieenzymatyczną?
Kataliza enzymatyczna opiera się na białkach lub kwasach nukleinowych o bardzo precyzyjnej strukturze trójwymiarowej. Tworzą one centrum aktywne, które rozpoznaje ściśle określony substrat i stabilizuje stan przejściowy, obniżając energię aktywacji. Katalizatory nieenzymatyczne, takie jak metale czy kwasy mineralne, zwykle działają mniej selektywnie i wymagają ostrzejszych warunków reakcji, np. wyższej temperatury, silniejszych odczynników czy innych rozpuszczalników.
Dlaczego enzymy są tak specyficzne wobec swoich substratów?
Specyficzność enzymów wynika z komplementarności przestrzennej i chemicznej centrum aktywnego względem substratu. Białko ma określoną geometrię kieszeni katalitycznej, rozmieszczenie ładunków oraz zdolność do tworzenia wiązań wodorowych i oddziaływań hydrofobowych. Tylko cząsteczka o odpowiednim kształcie i konfiguracji stereochemicznej może zostać prawidłowo związana i przekształcona. Dodatkowo wiele enzymów ulega dopasowaniu indukowanemu, co jeszcze bardziej wzmacnia selektywność.
Co wpływa na szybkość reakcji katalizowanych przez enzymy?
Na szybkość reakcji enzymatycznych wpływają głównie temperatura, pH, stężenie substratu oraz obecność aktywatorów lub inhibitorów. Każdy enzym ma swoje optimum temperaturowe i pH, przy którym jego struktura i jonizacja reszt katalitycznych są najkorzystniejsze. Wzrost stężenia substratu zwiększa szybkość do momentu wysycenia wszystkich centrów aktywnych. Dodatkowe czynniki, takie jak obecność jonów metali czy modyfikacje potranslacyjne, również mogą znacząco modyfikować aktywność katalityczną.
Czy enzymy zużywają się w trakcie reakcji chemicznej?
Enzymy nie ulegają trwałemu zużyciu w pojedynczym cyklu katalitycznym, ponieważ po przekształceniu substratu w produkt powracają do swojej wyjściowej formy. Ta regeneracja pozwala pojedynczej cząsteczce enzymu przeprowadzić ogromną liczbę cykli reakcji, zanim dojdzie do jej naturalnej degradacji w komórce. Jednak w praktyce enzymy mogą być inaktywowane przez wysoką temperaturę, skrajne pH, obecność inhibitorów nieodwracalnych, utleniaczy lub denaturujących rozpuszczalników, co skraca ich efektywny czas działania.
Jak wykorzystuje się katalizę enzymatyczną w przemyśle i medycynie?
W przemyśle enzymy stosuje się do przyspieszania i upraszczania procesów technologicznych, np. w produkcji żywności, detergentów, biopaliw czy związków farmaceutycznych. Umożliwiają one prowadzenie reakcji w łagodnych warunkach, z wysoką selektywnością i mniejszą ilością odpadów. W medycynie kataliza enzymatyczna jest celem terapii – wiele leków to inhibitory enzymów kluczowych dla patogenów lub szlaków chorobotwórczych. Enzymy służą też jako markery diagnostyczne i komponenty nowoczesnych biosensorów.