Potencjał błonowy jest jednym z kluczowych pojęć biologii komórki i fizjologii, a jednocześnie zjawiskiem, które łączy w sobie prawa chemii, fizyki i nauk o życiu. To właśnie dzięki różnicy ładunków elektrycznych po obu stronach błony komórkowej komórki nerwowe przewodzą impuls, mięśnie się kurczą, a komórki roślinne utrzymują odpowiednie napięcie turgorowe. Zrozumienie, czym jest potencjał błonowy, pozwala lepiej wyjaśnić nie tylko funkcjonowanie pojedynczej komórki, ale także działanie tkanek, narządów i całych organizmów.
Podstawy fizyczno-chemiczne potencjału błonowego
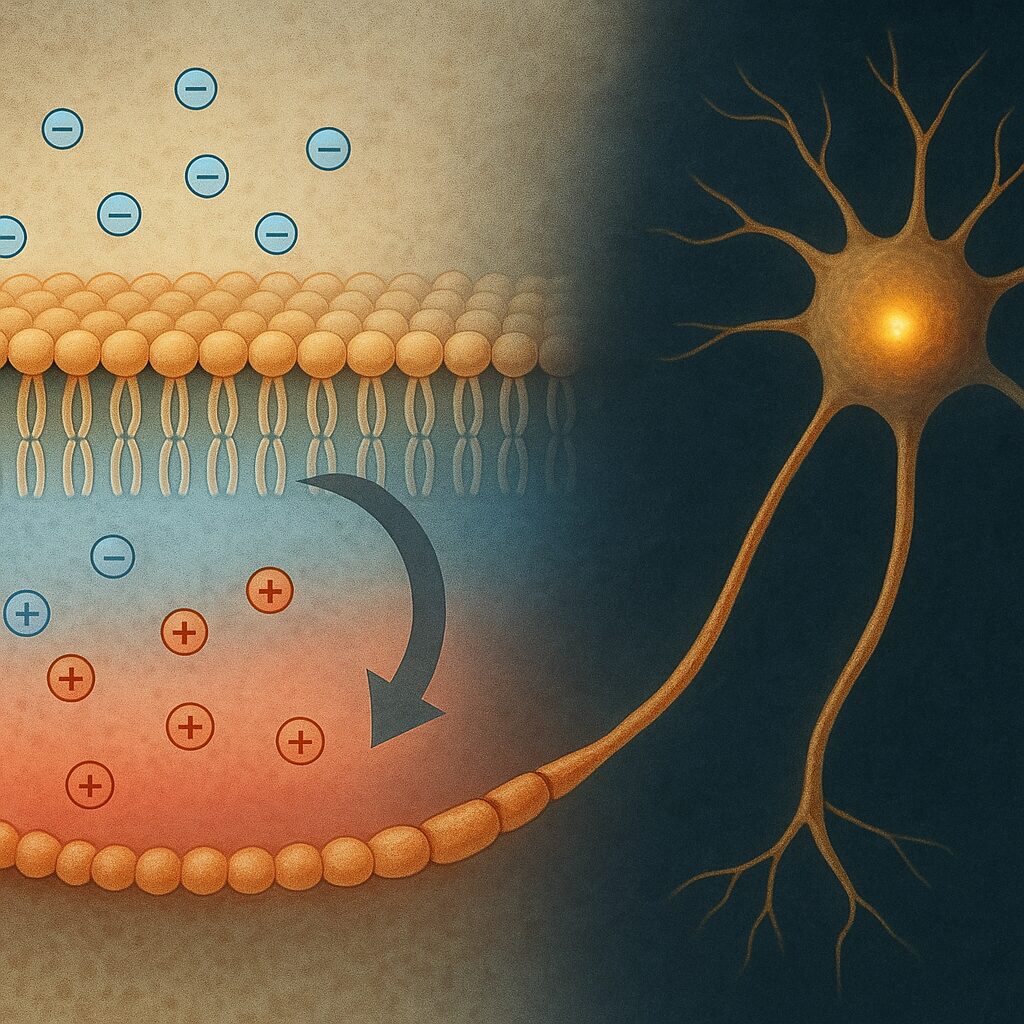
Błona komórkowa to cienka, dynamiczna struktura zbudowana głównie z podwójnej warstwy fosfolipidowej oraz zanurzonych w niej białek. Jej unikalną cechą jest selektywna przepuszczalność, dzięki której komórka może utrzymywać wewnętrzne środowisko w stanie odmiennym od otoczenia. Ta selektywność dotyczy w szczególności jonów, takich jak Na+, K+, Ca2+ czy Cl−, których rozmieszczenie po obu stronach błony jest ściśle kontrolowane.
Potencjał błonowy definiuje się jako różnicę potencjału elektrycznego między wnętrzem komórki a środowiskiem zewnętrznym, mierzony zazwyczaj w miliwoltach (mV). Konwencja przyjęta w biologii zakłada, że mierzony jest on jako potencjał wnętrza względem zewnątrz, dlatego potencjał spoczynkowy większości komórek ma wartość ujemną (np. −70 mV dla typowego neuronu). Oznacza to, że wnętrze komórki jest elektrycznie bardziej ujemne niż jej otoczenie.
Kluczową rolę w tworzeniu potencjału błonowego odgrywa zjawisko dyfuzji jonów oraz ich ruch w polu elektrycznym. Jony dążą do wyrównania gradientu stężeń (ruch z obszaru o większym stężeniu do mniejszego), ale jednocześnie ładunki elektryczne oddziałują ze sobą przyciągając się lub odpychając. W rezultacie ustala się dynamiczna równowaga, w której siła napędowa wynikająca z różnicy stężeń jest równoważona przez siłę elektryczną.
Potencjał równowagi dla pojedynczego jonu można obliczyć na podstawie równania Nernsta, które łączy wartości stężeń jonów po obu stronach błony z temperaturą i ładunkiem jonu. W praktyce potencjał błonowy jest wynikiem sumarycznego działania kilku jonów, dlatego bardziej złożone wzory, takie jak równanie Goldmana-Hodgkina-Katza, uwzględniają przepuszczalność błony dla różnych gatunków jonów. Niezależnie jednak od formalizmu matematycznego, istotą zjawiska jest nierównomierny rozkład jonów utrzymywany dzięki białkom błonowym i zróżnicowanej przepuszczalności błony.
Szczególne znaczenie ma tu aktywny transport jonów, przede wszystkim działanie pompy sodowo-potasowej (Na+/K+-ATPazy). Ten enzym błonowy, napędzany rozkładem ATP, usuwa z komórki jony sodu i wprowadza do niej jony potasu w stosunku 3:2. Ponieważ wypompowane są trzy dodatnie ładunki, a wprowadzane tylko dwa, pompa ta ma charakter elektrogeniczny i bezpośrednio przyczynia się do powstawania ujemnego potencjału wewnątrz komórki. Bez ciągłej pracy tego układu komórka stopniowo traciłaby zdolność utrzymania różnic stężeń i potencjału błonowego.
Potencjał spoczynkowy i potencjał czynnościowy
Najbardziej znanym przykładem funkcjonalnego znaczenia potencjału błonowego jest przewodzenie impulsów nerwowych. Neurony w stanie stabilnym nieprzekazującym sygnału utrzymują tzw. potencjał spoczynkowy. Jego typowa wartość wynosi około −60 do −80 mV, co odzwierciedla relatywnie dużą przepuszczalność błony dla jonów potasu w porównaniu z sodem. W warunkach spoczynku kanały potasowe o niskim stopniu otwarcia pozwalają na pewien wypływ K+ na zewnątrz, a jony ujemne zatrzymywane w komórce, takie jak aniony białczanowe, współtworzą przewagę ładunku ujemnego wewnątrz.
Kluczowe znaczenie ma tu różnica przewodności błony dla poszczególnych jonów. Dla jonów potasu jest ona stosunkowo duża, podczas gdy dla sodu relatywnie mała w stanie spoczynku. Dzięki temu potencjał spoczynkowy neuronu jest zbliżony do potencjału równowagi dla potasu. Jeżeli ta delikatna równowaga zostanie zakłócona, np. poprzez otwarcie dodatkowych kanałów sodowych, może dojść do gwałtownej depolaryzacji błony.
Depolaryzacja oznacza zmniejszenie różnicy potencjałów między wnętrzem a zewnętrzem komórki – przejście z wartości bardziej ujemnych w kierunku zera, a nawet dodatnich. W neuronie bodziec o odpowiedniej sile otwiera napięciowo zależne kanały sodowe, co prowadzi do napływu Na+ do komórki zgodnie z gradientem elektrochemicznym. W krótkim czasie wnętrze komórki staje się dodatnie względem otoczenia, co rejestrowane jest jako potencjał czynnościowy o wartości sięgającej około +30 mV.
Potencjał czynnościowy jest zjawiskiem o charakterze „wszystko albo nic”: jeżeli depolaryzacja osiągnie wartość progową, uruchamia się lawinowy mechanizm otwierania kolejnych kanałów sodowych. Po fazie szybkiej depolaryzacji następuje równie istotna repolaryzacja – zamykanie kanałów sodowych i otwieranie kanałów potasowych, co prowadzi do wypływu K+ i powrotu potencjału błonowego w okolice wartości spoczynkowej. Często obserwuje się również krótkotrwałą hiperpolaryzację, kiedy potencjał staje się bardziej ujemny niż w spoczynku, co wpływa na zdolność neuronu do generowania kolejnych impulsów.
Należy podkreślić, że propagacja potencjału czynnościowego wzdłuż aksonu nie polega na fizycznym „przemieszczaniu się” ładunku na duże odległości, lecz na lokalnych zmianach przepuszczalności błony komórkowej zachodzących sekwencyjnie w sąsiednich odcinkach. Oznacza to, że w każdym fragmencie błony dochodzi do przejściowego odwrócenia potencjału, które z kolei inicjuje podobną reakcję w następnym fragmencie. W przypadku włókien mielinowych impuls przeskakuje między przewężeniami Ranviera, co znacznie przyspiesza przewodzenie.
Podobne zasady, choć zwykle na innych skalach napięcia i czasu, obserwuje się w komórkach mięśniowych. W mięśniu sercowym potencjał błonowy odpowiada za generowanie rytmicznych pobudzeń oraz skoordynowane skurcze przedsionków i komór. Z kolei w mięśniach szkieletowych zmiany potencjału błonowego prowadzą do uwolnienia jonów wapnia z siateczki sarkoplazmatycznej, co umożliwia interakcję filamentów aktynowych i miozynowych. Bez kontrolowanej, odwracalnej zmiany potencjału błonowego precyzyjne sterowanie pracą mięśni byłoby niemożliwe.
Potencjał błonowy w różnych typach komórek
Choć neurony stanowią klasyczny model do studiowania potencjałów błonowych, niemal każda żywa komórka posiada różnicę potencjałów między wnętrzem a otoczeniem. Skala tego zjawiska oraz mechanizmy jego generowania mogą się jednak znacząco różnić w zależności od typu komórki i organizmu. Warto przyjrzeć się kilku przykładom pokazującym, jak zróżnicowany jest świat potencjałów błonowych w biologii.
W komórkach roślinnych błona plazmolemy otacza protoplast, ale dodatkowo obecna jest też ściana komórkowa oraz duże wakuole ograniczone tonoplastem. Potencjał błonowy roślin jest w znacznym stopniu kształtowany przez aktywność pomp protonowych (H+-ATPaz), które wypompowują jony wodorowe z cytoplazmy na zewnątrz. W efekcie po zewnętrznej stronie błony gromadzą się protony, tworząc gradient elektrochemiczny o dużej energii. Ten gradient wykorzystywany jest następnie do napędzania transportu innych jonów i cząsteczek, np. sacharozy czy azotanów, poprzez symporty i antyporty.
Tak zorganizowany system sprawia, że potencjał błonowy komórek roślinnych może osiągać wartości nawet poniżej −150 mV. Ma to bezpośrednie znaczenie dla procesów takich jak pobieranie składników mineralnych z gleby, regulacja otwierania i zamykania aparatów szparkowych, a także reakcje na bodźce świetlne czy dotykowe. Część roślin, jak mimoza wstydliwa, wykorzystuje gwałtowne zmiany potencjału błonowego do ruchów sejsmonastycznych – zginania liści i ogonków w odpowiedzi na mechaniczne drażnienie.
W komórkach zwierzęcych, poza neuronami i mięśniami, potencjał błonowy odgrywa istotną rolę m.in. w komórkach nabłonkowych, endokrynnych i układu odpornościowego. Komórki nabłonkowe wyściełające jelito, nerki czy drogi oddechowe wykorzystują różnice potencjału i gradienty jonowe do kierunkowego transportu substancji – wchłaniania składników odżywczych, wydzielania śluzu czy regulacji objętości płynów ustrojowych. Na przykład w kanaliku nerkowym przepływ jonów sodu i chlorków pod wpływem gradientów elektrochemicznych warunkuje proces resorpcji zwrotnej i utrzymania równowagi wodno-elektrolitowej.
Komórki wydzielania wewnętrznego, takie jak komórki beta trzustki, wykorzystują zmiany potencjału błonowego jako element przetwarzania informacji metabolicznej. Wzrost stężenia glukozy we krwi prowadzi do zwiększonego pobierania cukru przez te komórki, wzrostu ilości ATP i zamykania kanałów potasowych wrażliwych na ATP. W efekcie dochodzi do depolaryzacji błony, otwarcia napięciowo zależnych kanałów wapniowych i napływu Ca2+ do cytoplazmy. Podniesione stężenie wapnia wyzwala egzocytozę pęcherzyków zawierających insulinę, co pokazuje, że potencjał błonowy jest nie tylko zjawiskiem elektrycznym, ale i kluczowym elementem szlaku sygnałowego.
Istotną grupę stanowią również komórki układu odpornościowego – limfocyty, makrofagi, komórki tuczne. W ich przypadku potencjał błonowy wpływa na aktywację, proliferację oraz wydzielanie mediatorów zapalnych. Zmiany przewodnictwa dla jonów potasu i wapnia mogą modulować odpowiedź immunologiczną, a zaburzenia w pracy kanałów jonowych bywają związane z przewlekłymi stanami zapalnymi czy chorobami autoimmunologicznymi. Coraz więcej badań wskazuje także, że różnice potencjałów błonowych mogą regulować procesy proliferacji i różnicowania, w tym w komórkach nowotworowych, co otwiera nowe perspektywy dla terapii onkologicznych.
U mikroorganizmów, takich jak bakterie, potencjał błonowy jest częścią składową tzw. siły protonomotorycznej, czyli połączenia gradientu pH i różnicy potencjałów elektrycznych po obu stronach błony. Ten układ energetyczny napędza syntezę ATP poprzez syntazę ATP, transport aktywny wielu substancji, a także ruch bakterii za pomocą wici. W warunkach niedoboru energii, uszkodzenia błony lub działania antybiotyków, spadek potencjału błonowego jest jednym z wczesnych sygnałów kryzysu metabolicznego i może prowadzić do śmierci komórki.
Znaczenie potencjału błonowego dla homeostazy i zdrowia
Stabilny potencjał błonowy jest nieodzownym warunkiem zachowania homeostazy, czyli względnie stałych warunków wewnętrznych organizmu. Jego utrzymanie wymaga stałej współpracy kanałów jonowych, pomp, wymienników i transporterów, a każdy z tych elementów podlega precyzyjnej regulacji genetycznej i biochemicznej. Zaburzenia potencjału błonowego są często jednym z pierwszych objawów patologii na poziomie komórkowym, nawet zanim pojawią się widoczne zmiany morfologiczne.
W układzie nerwowym nieprawidłowości w funkcjonowaniu kanałów jonowych prowadzą do tzw. kanałopatii. Należą do nich różne postacie padaczki, migreny hemiplegiczne, niektóre arytmie serca oraz dziedziczne choroby mięśni. Mutacje w genach kodujących kanały sodowe, potasowe czy wapniowe mogą zaburzać generowanie i propagację potencjałów czynnościowych, prowadząc do nadmiernej pobudliwości lub przeciwnie – do osłabienia przewodzenia. Na poziomie komórkowym oznacza to zmianę kształtu i dynamiki potencjału błonowego, co bezpośrednio przekłada się na objawy kliniczne.
Szczególnie wrażliwy na odchylenia potencjału błonowego jest mięsień sercowy. Rytm pracy serca zależy od precyzyjnie skoordynowanych zmian potencjału w obrębie komórek węzła zatokowo-przedsionkowego, przedsionków, węzła przedsionkowo-komorowego oraz włókien Purkiniego. Leki antyarytmiczne, stosowane w terapii zaburzeń rytmu, działają głównie poprzez modulację przepływu jonów sodu, potasu i wapnia przez kanały błonowe. Poprzez wpływ na czas trwania i przebieg potencjału czynnościowego mogą zapobiegać niebezpiecznym migotaniom i częstoskurczom, ale jednocześnie ich niewłaściwe użycie niesie ryzyko wywołania innych, groźnych arytmii.
Równie ważną dziedziną, w której potencjał błonowy odgrywa dużą rolę, jest neurofarmakologia. Wiele leków wpływa pośrednio lub bezpośrednio na kanały jonowe i tym samym na pobudliwość neuronów. Leki znieczulające miejscowo blokują napięciowo zależne kanały sodowe, uniemożliwiając generowanie potencjałów czynnościowych w włóknach czuciowych i przewodzenie informacji bólowej. Z kolei niektóre leki przeciwpadaczkowe stabilizują inaktywowany stan kanałów sodowych lub modulują przepuszczalność dla potasu, zmniejszając nadmierną aktywność sieci neuronalnych.
Potencjał błonowy jest również ściśle związany z równowagą wodno-elektrolitową organizmu. Zmiany stężeń jonów w osoczu, takie jak hiperkaliemia czy hiponatremia, wpływają na wartości potencjału spoczynkowego komórek. Na przykład podwyższone stężenie potasu w płynach ustrojowych może depolaryzować błonę komórkową, zwiększając pobudliwość mięśnia sercowego i ryzyko arytmii. Odwrotnie, zbyt niskie stężenie potasu prowadzi do hiperpolaryzacji i utrudnia generowanie potencjałów czynnościowych, co objawia się osłabieniem mięśniowym i zaburzeniami przewodzenia nerwowego.
W kontekście badań nad nowotworami coraz większe zainteresowanie budzi rola potencjału błonowego w regulacji cyklu komórkowego i procesów różnicowania. Wiele komórek rakowych charakteryzuje się odmiennym potencjałem błonowym niż ich zdrowe odpowiedniki, co może wpływać na aktywność kanałów jonowych, szlaków sygnałowych i ekspresję genów. Pojawiają się koncepcje wykorzystywania modulacji potencjału błonowego jako uzupełniającej strategii terapeutycznej, zarówno poprzez leki wpływające na kanały jonowe, jak i dzięki fizycznym technikom oddziaływania na błonę komórkową.
Na poziomie bardziej ogólnym różnice potencjałów błonowych odgrywają istotną rolę w rozwoju organizmu. Podczas embriogenezy i regeneracji tkanek lokalne pola elektryczne mogą wpływać na migrację komórek, ich proliferację i wybór szlaków różnicowania. Badania w dziedzinie bioelektryczności rozwojowej sugerują, że gradienty potencjału błonowego stanowią swego rodzaju mapę informacyjną dla komórek, pomagając im „zdecydować”, w jaki typ tkanki się przekształcić i jak ukształtować strukturę rozwijającego się narządu.
Wreszcie, potencjał błonowy jest kluczowym narzędziem diagnostycznym i badawczym. Techniki takie jak patch-clamp pozwalają na rejestrację przepływu jonów przez pojedyncze kanały, co umożliwia szczegółowe poznanie ich właściwości i reakcji na leki. Z kolei barwniki wrażliwe na potencjał błonowy oraz elektrody mikroelektryczne umożliwiają analizę aktywności elektrycznej całych populacji komórek czy tkanek. Te narzędzia są szeroko stosowane zarówno w podstawowych badaniach biologicznych, jak i w testowaniu nowych substancji farmakologicznych.
Mechanizmy regulacji i plastyczność potencjału błonowego
Potencjał błonowy nie jest wartością sztywną; podlega dynamicznej regulacji w odpowiedzi na bodźce zewnętrzne i wewnętrzne. Jednym z najważniejszych mechanizmów jest modulacja liczby i właściwości kanałów jonowych obecnych w błonie. Komórka może zmieniać ekspresję genów kodujących te białka, wbudowywać je lub usuwać z błony poprzez procesy endo- i egzocytozy, a także modyfikować ich aktywność za pomocą fosforylacji, wiązania ligandów czy interakcji z innymi białkami.
W układzie nerwowym szczególne znaczenie ma zjawisko plastyczności elektrycznej. Neurony mogą dostosowywać swoją pobudliwość do długotrwałych zmian w aktywności sieci, np. w trakcie procesu uczenia się. Zwiększenie lub zmniejszenie liczby kanałów sodowych, potasowych czy wapniowych w określonych rejonach błony może zmieniać próg generowania potencjału czynnościowego, jego amplitudę, czas trwania oraz wzorce częstotliwości wyładowań. Ta zdolność adaptacji jest uzupełnieniem plastyczności synaptycznej i razem z nią tworzy podstawę dla pamięci i modyfikowalności zachowania.
Nie mniej istotna jest regulacja przez środowisko metaboliczne komórki. Stężenie ATP, poziom tlenu, obecność reaktywnych form tlenu, stan redoks czy aktywność kinaz i fosfataz wpływają na funkcjonowanie pomp i kanałów. Na przykład niedotlenienie powoduje spadek produkcji ATP, co może osłabiać działanie Na+/K+-ATPazy i stopniowo prowadzić do depolaryzacji błony. W mięśniu sercowym takie zmiany stanowią jeden z kluczowych mechanizmów uszkodzenia tkanki podczas zawału.
W komórkach roślinnych i mikroorganizmach warunki środowiskowe, takie jak zasolenie, pH, temperatura czy obecność toksyn, silnie oddziałują na potencjał błonowy. Rośliny narażone na nadmiar soli w glebie muszą modyfikować transport jonów sodu i potasu, aby utrzymać odpowiednie wartości potencjału i zachować funkcjonalność komórek korzeniowych. Bakterie z kolei reagują na środki przeciwdrobnoustrojowe często poprzez zmiany w przepuszczalności błony i reorganizację systemów transportu jonów, co pomaga im przetrwać niekorzystne warunki.
Z perspektywy inżynierii biomedycznej i biotechnologii umiejętne manipulowanie potencjałem błonowym staje się narzędziem do sterowania zachowaniem komórek. W badaniach nad komórkami macierzystymi wykazano, że zmiana ich potencjału błonowego – np. poprzez farmakologiczną modulację kanałów jonowych – może wpływać na decyzję o pozostaniu w stanie niezróżnicowanym lub wejściu na ścieżkę różnicowania w kierunku określonego typu tkanki. Otwiera to możliwość projektowania terapii regeneracyjnych opartych nie tylko na czynnikach wzrostu i sygnałach biochemicznych, ale także na kontrolowanych bodźcach bioelektrycznych.
Współczesne badania nad potencjałem błonowym obejmują też rozwój nowoczesnych narzędzi pomiarowych i interwencyjnych. Elektrody wielokanałowe o wysokiej gęstości pozwalają jednocześnie rejestrować aktywność setek czy tysięcy neuronów, co umożliwia analizę złożonych sieci neuronalnych. Z kolei optogenetyka, wykorzystująca białka światłoczułe pełniące funkcję kanałów jonowych, pozwala precyzyjnie sterować potencjałem błonowym wybranych komórek za pomocą światła. Dzięki temu możliwe jest badanie związków między aktywnością elektryczną a zachowaniem, a także opracowywanie nowych strategii terapeutycznych w chorobach neurologicznych.
FAQ – najczęściej zadawane pytania
Czym różni się potencjał spoczynkowy od potencjału czynnościowego?
Potencjał spoczynkowy to stała, ujemna różnica potencjałów między wnętrzem komórki a jej otoczeniem w stanie, gdy komórka nie przekazuje sygnału. Utrzymują go głównie pompa sodowo-potasowa i selektywna przepuszczalność błony dla jonów potasu. Potencjał czynnościowy występuje, gdy komórka – np. neuron lub komórka mięśniowa – zostaje pobudzona bodźcem progowym i dochodzi do szybkiej, przejściowej depolaryzacji oraz następującej po niej repolaryzacji błony.
Dlaczego wnętrze komórki jest zwykle naładowane ujemnie?
Ujemny ładunek wnętrza komórki wynika głównie z nierównomiernego rozmieszczenia jonów i obecności dużych, ujemnie naładowanych cząsteczek, takich jak białka i aniony organiczne, które nie mogą swobodnie przenikać przez błonę. Błona komórkowa jest bardziej przepuszczalna dla jonów potasu niż dla sodu, co sprzyja wypływowi K+ na zewnątrz. Jednocześnie pompa Na+/K+ usuwa z komórki więcej dodatnich ładunków niż do niej wprowadza, co dodatkowo potęguje ujemny potencjał błonowy.
Jak mierzy się potencjał błonowy w praktyce laboratoryjnej?
Do pomiaru potencjału błonowego stosuje się mikroelektrody szklane wprowadzane bezpośrednio do komórki lub technikę patch-clamp, w której szklana pipeta tworzy szczelne połączenie z fragmentem błony. Rejestrowana jest różnica napięcia między wnętrzem komórki a elektrodą odniesienia w płynie zewnątrzkomórkowym. Coraz częściej używa się też barwników fluorescencyjnych wrażliwych na potencjał, pozwalających monitorować zmiany w czasie rzeczywistym w wielu komórkach jednocześnie, co jest przydatne przy badaniu tkanek i sieci neuronalnych.
Jakie czynniki mogą zaburzać potencjał błonowy komórki?
Na potencjał błonowy wpływają przede wszystkim stężenia jonów w środowisku, sprawność pomp jonowych oraz funkcjonowanie kanałów w błonie. Zaburzenia elektrolitowe, jak hiperkaliemia czy hiponatremia, zmieniają gradienty stężeń i powodują depolaryzację lub hiperpolaryzację. Toksyczne substancje, leki, niedotlenienie lub uszkodzenia metaboliczne mogą hamować Na+/K+-ATPazę i inne transportery. Z kolei mutacje w genach kanałów jonowych prowadzą do kanałopatii, objawiających się m.in. arytmiami serca czy napadami padaczkowymi.
Dlaczego potencjał błonowy jest tak ważny dla funkcjonowania organizmu?
Potencjał błonowy jest podstawą przekazywania informacji elektrycznej w układzie nerwowym, skurczu mięśni, wydzielania hormonów oraz wielu procesów transportu przez błonę komórkową. Dzięki niemu neurony mogą szybko przesyłać impulsy, serce utrzymuje rytmiczne bicie, a komórki wydzielnicze reagują na zmiany środowiska. Utrata zdolności do generowania i utrzymywania prawidłowego potencjału błonowego prowadzi do ciężkich zaburzeń, od porażeń mięśniowych, przez arytmie, po śmierć komórki, dlatego jego regulacja jest kluczowa dla zdrowia.

