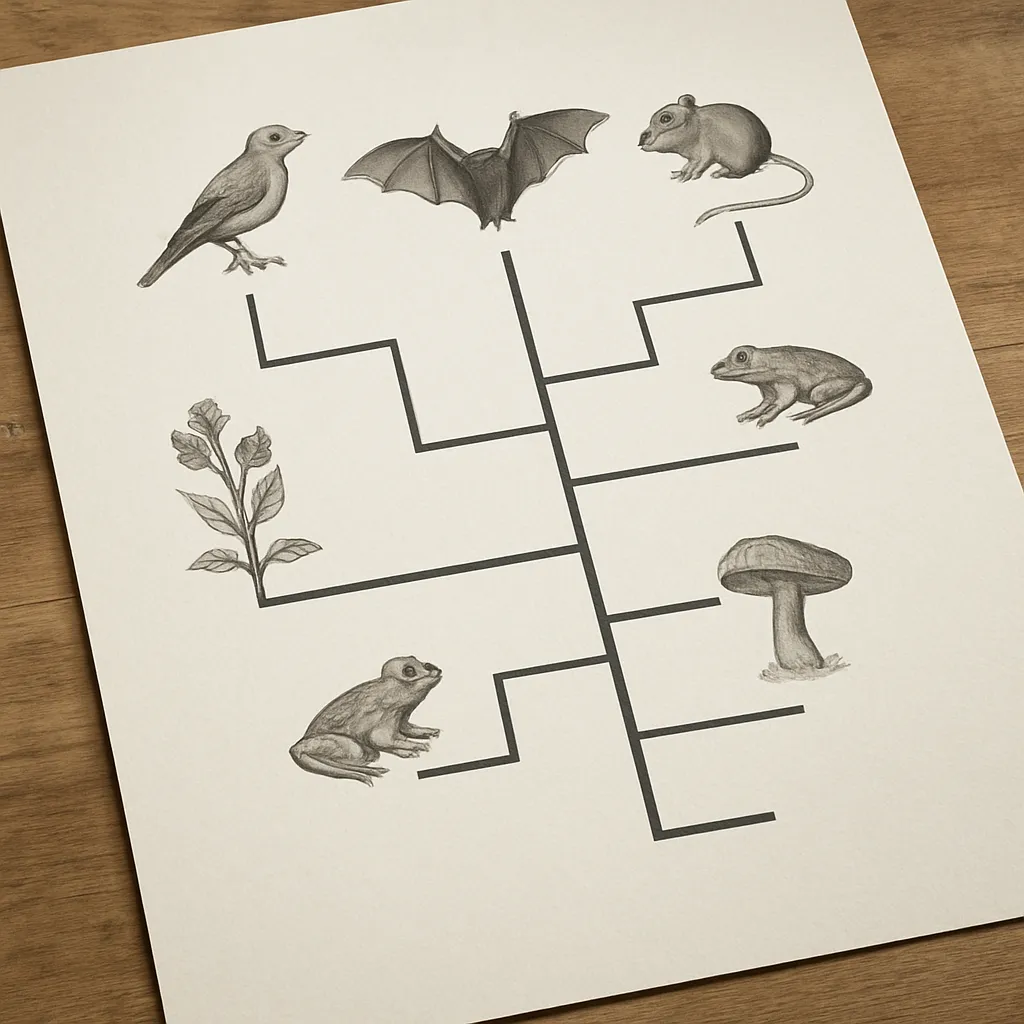
Drzewa filogenetyczne to jedno z najważniejszych narzędzi współczesnej biologii, pozwalające zrozumieć pokrewieństwo między organizmami oraz kierunki, w jakich przebiegała ewolucja życia na Ziemi. Dzięki nim można odtworzyć historię przemian dziedziczonych cech, śledzić losy całych linii ewolucyjnych oraz łączyć dane z genetyki, morfologii, ekologii czy paleontologii w spójną całość. Poznanie zasad tworzenia i interpretowania drzew filogenetycznych jest kluczowe, by właściwie rozumieć współczesną systematykę organizmów i sens pojęcia wspólnego przodka.
Podstawy pojęcia drzewa filogenetycznego
Drzewo filogenetyczne to graficzne przedstawienie hipotezy o ewolucyjnych relacjach między gatunkami, genami lub innymi jednostkami biologicznymi. Zazwyczaj rysuje się je w formie rozgałęziającej się struktury przypominającej drzewo, w której linie (gałęzie) odzwierciedlają ciągłość dziedziczenia, a węzły rozgałęzień oznaczają punkty rozdzielenia się linii, czyli zdarzenia specjacji lub duplikacji genów.
Kluczowe jest zrozumienie, że takie drzewo jest zawsze modelem, a nie dosłownym zapisem historii. Oznacza to, że bazuje na danych, które posiadamy w danym momencie, oraz na metodach analizy, które stosujemy. Wraz z pojawianiem się nowych informacji drzewo może być modyfikowane: zmienia się pozycja niektórych gatunków, długość gałęzi, a czasem także sama liczba rozgałęzień. Biolog nigdy nie mówi, że drzewo filogenetyczne jest ostateczną prawdą, lecz że jest to najlepiej wspierana dowodami propozycja przebiegu ewolucji w danej grupie organizmów.
Na drzewie filogenetycznym znajdują się najczęściej trzy główne elementy: liście (końce gałęzi), linie (gałęzie) oraz węzły. Liście reprezentują współczesne organizmy albo sekwencje genetyczne, które analizujemy. Gałęzie to ścieżki ewolucyjne, wzdłuż których zachodzą mutacje, zmiany cech i adaptacje. Węzły oznaczają hipotetycznych wspólnych przodków, których nie obserwujemy bezpośrednio, ale których istnienie wynika z układu pokrewieństw. Często dodatkowo wyróżnia się korzeń drzewa, czyli punkt odpowiadający najdalszemu, najstarszemu przodkowi danej grupy.
Drzewo filogenetyczne można tworzyć dla bardzo różnych poziomów organizacji życia. Może ono przedstawiać pokrewieństwo pojedynczych genów w genomie, relacje między szczepami jednego gatunku, pozycje całych rodzin, rzędów czy gromad, aż po filogenetyczne drzewo całego życia, obejmujące bakterie, archeony oraz eukarionty. Zasada pozostaje ta sama: linie rozchodzą się tam, gdzie w przeszłości zaszło zdarzenie rozdzielenia się populacji lub sekwencji na dwie niezależne ścieżki dziedziczenia.
Istotne jest odróżnienie drzewa filogenetycznego od prostego diagramu podobieństwa. Dwa gatunki mogą być do siebie morfologicznie podobne, lecz wcale nie muszą być najbliżej spokrewnione. Drzewo ma przedstawiać historię genealogiczną, a nie wyłącznie stopień podobieństwa zewnętrznego. Zjawiska takie jak konwergencja ewolucyjna (niezależne wykształcenie podobnych cech w niespokrewnionych liniach) powodują, że podobieństwo fenotypu bywa mylące. Właśnie dlatego współczesne filogenetyki tak silnie opierają się na danych molekularnych.
Elementy i interpretacja drzew filogenetycznych
Rozumienie, co oznaczają poszczególne części drzewa filogenetycznego, jest niezbędne do prawidłowej interpretacji wyników badań. W podstawowej formie wyróżniamy drzewa zakorzenione i niezakorzenione. Drzewo zakorzenione ma jasno wskazany kierunek czasu: od korzenia (najstarszego przodka) do liści (najmłodsze współczesne jednostki). Drzewo niezakorzenione pokazuje tylko wzajemne relacje, ale nie określa, który węzeł jest najstarszy – nie znamy wtedy kierunku ewolucji, jedynie sieć pokrewieństw.
Na drzewie zakorzenionym można odczytać kolejność rozdzielania się linii. Jeśli przyjrzymy się konkretnemu węzłowi, w którym gałąź dzieli się na dwie, rozróżniamy grupę siostrzaną (clade sister) i grupę wyjściową (outgroup). Grupy siostrzane są swoimi najbliższymi krewnymi, to znaczy, że mają siebie nawzajem za najbliższych wspólnych przodków. Outgroup to natomiast linia, którą traktujemy jako punkt odniesienia przy określaniu kierunku ewolucji cech w analizowanej grupie.
Istotne pojęcie stanowi klad (grupa monofiletyczna). Jest to zbiór organizmów obejmujący wspólnego przodka i wszystkich jego potomków. Klad można „odciąć” jednym cięciem gałęzi drzewa – wszystko, co znajdzie się po jednej stronie cięcia, to kompletny klad. Biolodzy starają się, aby współczesna systematyka odzwierciedlała właśnie takie klady. Unika się za to grup parafiletycznych (obejmujących przodka, ale nie wszystkie linie potomne) oraz polifiletycznych (zbierających razem niespokrewnione linie tylko dlatego, że są do siebie podobne).
Przykładem historycznego problemu była tradycyjna grupa „ryby”. W ujęciu filogenetycznym wyłączenie z niej czworonogów sprawia, że jest to grupa parafiletyczna, ponieważ wspólny przodek wszystkich „ryb” jest jednocześnie przodkiem zwierząt lądowych. Nowoczesne ujęcie woli więc mówić o kladzie zwanym kręgowce żuchwowe i dalej wydzielać w nim linie, które są kladami, zamiast używać pojęć sprzecznych z drzewem filogenetycznym.
Drzewo może być skalowane czasem lub liczbą zmian. W przypadku drzew skalowanych czasowo długość gałęzi odpowiada wiekowi linii – dłuższa gałąź oznacza więcej czasu, który upłynął od rozdzielenia się od wspólnego przodka. Z kolei w drzewach skalowanych liczbą zmian długość gałęzi reprezentuje liczbę substytucji nukleotydowych czy aminokwasowych. Takie podejście stosuje się w filogenetyce molekularnej, gdzie kluczowe jest oszacowanie tempo mutacji w sekwencjach DNA czy białek.
Warto zwrócić uwagę, że sposób rysowania drzewa – poziomo lub pionowo, z gałęziami prostymi lub łukowymi – nie ma znaczenia dla interpretacji. Istotne są relacje rozgałęzień i położenie węzłów względem siebie, nie zaś dokładny układ graficzny. Z tego powodu to samo drzewo można narysować na wiele sposobów, które na pierwszy rzut oka wyglądają różnie, ale wyrażają tę samą strukturę pokrewieństw.
Kluczowe umiejętności przy analizie drzewa to:
- odczytywanie, które gatunki są najbliżej spokrewnione (wspólny węzeł najpóźniej w czasie),
- rozpoznawanie kladów obejmujących przodka i wszystkich potomków,
- rozumienie znaczenia długości gałęzi, jeśli drzewo jest skalowane,
- interpretacja wartości wsparcia statystycznego (np. bootstrap),
- rozróżnianie hipotez ugruntowanych od słabiej popartych przez dane.
W drzewach filogenetycznych często pojawiają się także polytomie – węzły, z których wychodzi więcej niż dwie gałęzie. Mogą one oznaczać prawdziwe, niemal równoczesne rozdzielenie się kilku linii, ale najczęściej odzwierciedlają brak rozstrzygających danych i niepewność co do kolejności zdarzeń. Dlatego interpretując drzewo, należy zawsze pamiętać o ograniczeniach metody i jakości użytego materiału.
Metody tworzenia drzew filogenetycznych
Budowanie drzewa filogenetycznego polega na przełożeniu danych biologicznych – cech morfologicznych, sekwencji genetycznych, struktur białek czy informacji paleontologicznych – na uporządkowaną, matematyczną reprezentację pokrewieństwa. Zasadniczym etapem jest wybór danych, przygotowanie ich do analizy, zastosowanie odpowiedniej metody inferencji filogenetycznej oraz ocena wiarygodności uzyskanego drzewa.
W przypadku danych molekularnych pierwszym krokiem jest zebranie sekwencji DNA, RNA lub białek od organizmów, które chcemy porównać. Sekwencje muszą reprezentować homologiczne regiony – oznacza to, że pochodzą od wspólnego przodka i pełnią w przybliżeniu tę samą funkcję. Następnie wykonuje się wyrównanie sekwencji (multiple sequence alignment), czyli takie ich zestawienie, aby pozycje odpowiadające tym samym miejscom w cząsteczce znalazły się w tych samych kolumnach.
Na tym etapie szczególnie ważna jest identyfikacja pozycji informatywnych, czyli takich, w których różnice między sekwencjami mogą pomóc w rozstrzygnięciu przebiegu ewolucji. Zbyt zmienne regiony bywają nasycone mutacjami i trudne do interpretacji, natomiast całkowicie zachowane fragmenty nie wnoszą informacji o relacjach wewnątrz grupy. Z tego powodu filogenetycy dobierają markery molekularne o odpowiednim tempie ewolucji do skali czasowej, którą chcą badać.
Klasyczne metody konstrukcji drzew można podzielić na kilka kategorii. Metody odległościowe, takie jak UPGMA czy Neighbor-Joining, opierają się na macierzy odległości między sekwencjami (np. liczbie różnic nukleotydowych skorygowanej odpowiednim modelem substytucji). Na podstawie tych odległości algorytm stopniowo łączy najbliższe pary sekwencji i rekonstruuje drzewo, które najlepiej odtwarza wzór obserwowanych dystansów.
Metody parsymonii minimalizują łączną liczbę zmian niezbędnych do wyjaśnienia obserwowanych różnic między sekwencjami lub cechami morfologicznymi. Zgodnie z zasadą brzytwy Ockhama preferuje się scenariusze najprostsze, zakładając, że ewolucja nie wprowadza nadmiarowych przekształceń, jeśli nie jest to konieczne. Jednak rzeczywista ewolucja nie zawsze jest „oszczędna”, a skomplikowane zjawiska, takie jak ewolucja mozaikowa czy homoplazje, mogą sprawić, że rozwiązanie parsymoniczne nie będzie prawidłowe.
Najbardziej zaawansowane podejścia wykorzystują metody statystyczne, w tym maksymalne prawdopodobieństwo (maximum likelihood) i metody bayesowskie. W tym ujęciu definiuje się model ewolucji sekwencji (np. różne prawdopodobieństwa substytucji poszczególnych nukleotydów, zróżnicowane tempo zmian w różnych pozycjach), a następnie oblicza, które drzewo najlepiej tłumaczy obserwowane dane, tzn. przy którym prawdopodobieństwo zaobserwowania danej macierzy sekwencji jest największe.
Metody bayesowskie idą krok dalej, pozwalając na obliczenie pełnego rozkładu prawdopodobieństwa drzew, a nie tylko wskazanie jednego najlepszego drzewa. Wykorzystuje się tu łańcuchy Markowa Monte Carlo (MCMC), aby próbować przestrzeń wszystkich możliwych drzew. Wynikiem takiej analizy są zwykle drzewo konsensusowe z wartością wsparcia bayesowskiego dla kolejnych węzłów, informujące, jak często dana relacja pojawiała się w próbkach ze wspomnianego rozkładu.
Ocenę wiarygodności drzewa przeprowadza się m.in. za pomocą procedury bootstrap. Polega ona na wielokrotnym, losowym próbkowaniu kolumn wyrównania sekwencji z powtórzeniami i rekonstrukcji drzewa dla każdej próbki. Następnie oblicza się, jak często dany węzeł pojawia się w wynikach. Wysoka wartość bootstrap (np. powyżej 90%) wskazuje, że dana relacja jest stabilna wobec losowych wahań w danych, choć nie gwarantuje absolutnej poprawności.
Kolejnym wyzwaniem w filogenetyce jest uwzględnienie zjawisk utrudniających prostą, „drzewiastą” reprezentację ewolucji. Wśród nich wymienia się poziomy transfer genów, hybrydyzację między gatunkami, duplikacje i utraty genów oraz rekombinację. Takie zjawiska mogą powodować, że różne geny opowiadają odmienne „historie” ewolucyjne, a jedno drzewo gatunkowe nie wystarcza do opisania pełnego obrazu. W odpowiedzi na te problemy rozwija się koncepcja sieci filogenetycznych oraz złożonych modeli łączących drzewa gatunkowe z drzewami genowymi.
Zastosowania drzew filogenetycznych w nauce
Drzewa filogenetyczne są fundamentem współczesnej taksonomii i systematyki. Zastępują tradycyjne klasyfikacje oparte wyłącznie na cechach morfologicznych, które często nie oddawały rzeczywistego pokrewieństwa. Dzięki drzewom możliwe jest tworzenie systemów nazw zgodnych z zasadą monofiletyczności, tak aby każda nazwana jednostka (rodzina, rząd, gromada) odzwierciedlała rzeczywisty klad. Pozwala to uporządkować ogromną różnorodność życia w sposób zgodny z jego historią ewolucyjną.
Jednym z najważniejszych zastosowań drzew filogenetycznych jest badanie pochodzenia i rozprzestrzeniania się chorób zakaźnych. Analizując sekwencje wirusów lub bakterii z różnych miejsc i okresów, można odtworzyć drzewo ich pokrewieństwa i zrekonstruować drogę transmisji. Tego typu analizy wykorzystywano przy śledzeniu epidemii HIV, grypy, Ebola czy SARS-CoV-2. Drzewo filogenetyczne pozwala zidentyfikować źródła zakażeń, momenty skoków międzygatunkowych oraz tempo rozprzestrzeniania się nowych wariantów patogenu.
W biologii ewolucyjnej drzewa filogenetyczne służą do badania dynamiki powstawania gatunków i wymierań. Analizując kształt drzewa (np. liczbę i długość gałęzi, rozkład czasu rozgałęzień), można wnioskować o tempie specjacji, istnieniu radiacji adaptacyjnych czy wpływie zmian środowiskowych na różnorodność grupy. Modele makroewolucyjne łączą dane filogenetyczne z informacjami paleontologicznymi, aby tworzyć ilościowe opisy historii życia na Ziemi.
Drzewa są również niezbędne do rekonstrukcji ewolucji cech fenotypowych. Łącząc informacje o charakterystyce współczesnych gatunków (np. kształcie ciała, sposobie odżywiania, typie rozrodu) z drzewem pokrewieństwa, można wnioskować, jakie właściwości miał przodek danej grupy i ile razy dana cecha powstała niezależnie. Analizy tego typu ujawniły m.in. wielokrotne, niezależne pojawianie się jaskrawego ubarwienia ostrzegawczego, złożonych systemów opieki nad potomstwem czy zjawiska ewolucji konwergentnej w zupełnie różnych liniach organizmów.
W filogeografii łączy się drzewa filogenetyczne z danymi przestrzennymi. Celem jest zrozumienie, jak linie ewolucyjne rozprzestrzeniały się w czasie i przestrzeni, jakie bariery geograficzne wpływały na rozdzielanie populacji i jak zmiany klimatu kształtowały obecną dystrybucję gatunków. Dzięki takim badaniom lepiej poznajemy, jak powstały endemiczne fauny wysp, jakie trasy migracyjne wybierały dawne populacje oraz jak przeszłe zlodowacenia wpływały na rozmieszczenie roślin i zwierząt.
Nie można pominąć roli drzew filogenetycznych w medycynie i farmakologii. Analiza ewolucyjnych relacji między białkami pozwala przewidywać funkcje nieopisanych jeszcze genów, identyfikować potencjalne cele dla leków oraz zrozumieć mechanizmy oporności na antybiotyki i chemioterapeutyki. Porównując drzewa białek ludzkich i patogennych, można wykrywać przypadki horyzontalnego transferu genów lub wspólnego pochodzenia szlaków metabolicznych, co dostarcza wskazówek dla projektowania terapii.
Drzewa filogenetyczne odgrywają także istotną rolę w badaniach nad pochodzeniem człowieka. Analiza sekwencji mitochondrialnego DNA, chromosomu Y i genomu jądrowego pozwoliła odtworzyć historię migracji Homo sapiens, relacje z neandertalczykami i denisowianami oraz złożony proces mieszania się populacji na przestrzeni tysięcy lat. Te same metody stosuje się w genetyce populacyjnej zwierząt hodowlanych i dzikich, aby śledzić przepływ genów, identyfikować wąskie gardła demograficzne i planować strategie ochrony zagrożonych gatunków.
Wreszcie, rozwój genomiki porównawczej uczynił drzewa filogenetyczne nieodzownym narzędziem przy interpretacji całych genomów. Porównując ułożenie genów, struktury chromosomów i konserwowane regiony regulacyjne w świetle drzewa pokrewieństwa, można identyfikować dawne duplikacje genomowe, translokacje i inwersje, a także śledzić powstawanie nowych funkcji biologicznych. Ewolucyjny kontekst zapobiega błędnym wnioskom opartym na prostym zestawieniu sekwencji bez uwzględnienia ich historii.
Ograniczenia i wyzwania w interpretacji filogenezy
Mimo ogromnej użyteczności drzewa filogenetyczne niosą ze sobą istotne ograniczenia. Po pierwsze, są to zawsze hipotezy – potencjalnie poprawne wyjaśnienia historii ewolucyjnej, ale podatne na rewizje. Zmiana zestawu analizowanych gatunków, wybór innego modelu ewolucji sekwencji czy uwzględnienie dodatkowych danych paleontologicznych może istotnie zmodyfikować kształt drzewa. Nauka akceptuje tę otwartość, postrzegając filogenezę jako dziedzinę, w której wiedza stale się aktualizuje wraz z pojawianiem się nowych linii dowodowych.
Po drugie, dane molekularne obarczone są zjawiskami, które zaciemniają sygnał ewolucyjny. Do najważniejszych należą homoplazje, czyli zbieżne lub równoległe mutacje, w wyniku których różne linie osiągają podobne stany sekwencji niezależnie od wspólnego pochodzenia. Przy wysokich szybkościach substytucji może dochodzić do saturacji – wielokrotne zmiany w tej samej pozycji maskują wcześniejsze zdarzenia. W takich sytuacjach proste miary odległości sekwencyjnej stają się mylące, a bardziej złożone modele tylko częściowo rozwiązują problem.
Po trzecie, historia genów nie zawsze jest identyczna z historią gatunków. Zjawiska takie jak niekompletne sortowanie rodowodów (incomplete lineage sorting), duplikacje i utraty genów czy wspomniany już poziomy transfer genów prowadzą do rozbieżności między drzewem genowym a drzewem gatunkowym. W skrajnych przypadkach różne fragmenty genomu sugerują wzajemnie sprzeczne scenariusze pokrewieństwa. Wymaga to stosowania metod, które jawnie modelują relacje między wieloma drzewami genowymi i nadrzędnym drzewem gatunkowym.
Po czwarte, wiele metod filogenetycznych zakłada względną jednorodność procesów ewolucyjnych w czasie i w różnych liniach, co rzadko jest w pełni prawdziwe. Tempo mutacji może zmieniać się wskutek różnic w wielkości populacji, trybie życia, mechanizmach naprawy DNA czy poziomie stresu środowiskowego. Ignorowanie tych czynników może prowadzić do błędnych oszacowań długości gałęzi i czasu rozdzielenia linii. W odpowiedzi rozwijane są tzw. relaksowane zegary molekularne, dopuszczające zróżnicowanie tempa zmian w różnych częściach drzewa.
Po piąte, interpretacja drzew filogenetycznych w kontekście ewolucji cech wymaga ostrożności. Rekonstrukcje stanów przodków, liczby niezależnych powstawań danego przystosowania czy scenariuszy transformacji często opierają się na modelach o uproszczonych założeniach. Wiele cech ma złożone podłoże genetyczne i rozwojowe, a to oznacza, że proste modele zmian stanów (np. 0/1 dla obecności czy braku cechy) mogą zbyt mocno uogólniać skomplikowaną rzeczywistość biologiczną.
Ostatnim, ale istotnym wyzwaniem jest komunikacja wyników filogenetycznych poza środowiskiem specjalistów. Graficzna forma drzew sprzyja uproszczeniom i złym interpretacjom, takim jak traktowanie organizmów „położonych wyżej” jako bardziej zaawansowanych czy ewolucyjnie „lepszych”. Tymczasem każde współcześnie istniejące gatunki są w tym samym stopniu wynikiem długiej historii ewolucji, a kategorie postępu i wyższości są w biologii ewolucyjnej pojęciami mylnymi. Drzewo pokazuje rozgałęzienia linii, a nie hierarchię wartości.
FAQ
Czym dokładnie różni się drzewo filogenetyczne od klasycznej klasyfikacji gatunków?
Klasyczna, dawna klasyfikacja opierała się głównie na zewnętrznym podobieństwie organizmów, często ignorując rzeczywiste powiązania genealogiczne. Drzewo filogenetyczne przedstawia hipotezę o historii pochodzenia gatunków, bazując na danych molekularnych, morfologii i skamieniałościach. Zamiast grupować organizmy „podobne do siebie”, łączy te, które dzielą wspólnego przodka. Dzięki temu współczesna systematyka dąży do tworzenia kladów, a nie sztucznych, parafiletycznych zgrupowań.
Skąd wiemy, że dane drzewo filogenetyczne jest wiarygodne?
Wiarygodność drzewa ocenia się, analizując m.in. jakość danych (długość i zmienność sekwencji, dobór markerów), spójność wyników z różnych genów i metod, wartości wsparcia statystycznego (np. bootstrap, wsparcie bayesowskie) oraz zgodność z niezależnymi liniami dowodów, jak dane paleontologiczne czy biogeograficzne. Jedno drzewo nie jest niepodważalną prawdą, lecz najlepszą obecną hipotezą. Im więcej zbieżnych źródeł wskazuje na podobny kształt drzewa, tym większe zaufanie budzi dana rekonstrukcja.
Czy drzewo filogenetyczne pokazuje „postęp” ewolucyjny?
Drzewo filogenetyczne nie opisuje postępu ani hierarchii gatunków, a jedynie ich pokrewieństwo i kolejność rozgałęzień linii. Wszystkie współcześnie żyjące organizmy są potomkami równie długiej historii ewolucyjnej, choć różnią się zakresem zmian. Położenie na drzewie nie oznacza wyższości czy „bardziej zaawansowanego” stadium rozwoju. Tego typu interpretacje wynikają z antropocentrycznego spojrzenia. Biolodzy używają drzew do badania adaptacji i specjacji, a nie do wartościowania gatunków.
Dlaczego różne geny czasem sugerują inne drzewa dla tych samych gatunków?
Różne geny mogą mieć odmienne historie, ponieważ podlegają zjawiskom takim jak niekompletne sortowanie rodowodów, duplikacje i utraty, rekombinacja czy poziomy transfer genów. Gen może „pamiętać” wydarzenia z innej skali czasowej lub być podatny na inne siły selekcyjne niż reszta genomu. W efekcie pojedyncze drzewo genowe nie zawsze odzwierciedla drzewa gatunkowego. Dlatego nowoczesna filogenetyka łączy informacje z wielu genów i stosuje modele, które jawnie rozróżniają filogenezę genów i gatunków.
W jaki sposób drzewa filogenetyczne pomagają śledzić rozwój epidemii?
Analizując sekwencje patogenu (np. wirusa) z różnych miejsc i czasów, można zrekonstruować drzewo ich pokrewieństwa. Układ gałęzi i długości odzwierciedla rozgałęzienia linii zakażeń i tempo gromadzenia mutacji. Dzięki temu da się wnioskować o źródłach wybuchu epidemii, identyfikować superrozkazicieli, określać moment skoku międzygatunkowego oraz śledzić rozprzestrzenianie się konkretnych wariantów. Informacje te wspierają planowanie strategii zdrowia publicznego i ocenę skuteczności interwencji.
